PTC Tasting Lab (Parts 1 & 2)
advertisement
")
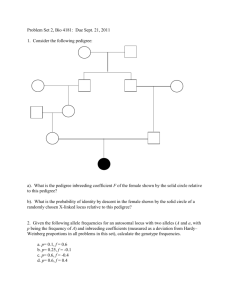
Hardy-Weinberg PTC Tasting Lab Purpose: Calculate allele, genotype and phenotype frequencies for two alleles of a gene, using the Hardy-Weinberg equation Procedure: Part 1: Estimation of Gene Frequencies for the Trait to Taste PTC Within a Small Sample Population This experiment deals with the determination of the gene frequency of a human trait amongst students with no known selective advantage. The ability to taste the chemical phenylthiocarbamide, PTC, is one such human trait. The ability to taste PTC is due to the presence of a dominant allele, T. Therefore, all tasters will either be homozygous, TT, or heterozygous, Tt. Non-tasters will be homozygous for the recessive gene, tt. Using the Hardy-Weinberg equation, first determine the frequency of q2, which corresponds the homozygous recessive gene t, which is directly determined from your class data. 1. 2. 3. 4. 5. Student groups should obtain a PTC taste strip and a control strip. Every member of the group should first taste the control strip of paper. Every person should now taste the PTC strip of paper. Compare the taste of the control and the PTC paper. If you are a PTC taster, the strip will taste SUPER BITTER. Non-tasters will not notice a difference between the control strip and the PTC strip. For the class, record the total number of tasters and the total number of non-tasters in your group on the whiteboard. Record the class population data in the table below and determine the frequency of p and q in both the class population and the north American population. Table 1: Phenotypes and Gene Frequencies for Trait to Taste PTC. Part 1 Analysis: 1. What is the frequency of homozygous tasters in our class? 2. What is the frequency of heterozygous tasters in our class? 3. What is the frequency of homozygous non-tasters in our class? 4. Determine the percentages of the three genotypes for this trait in our class. PART 2: Testing an Idealized Hardy-Weinberg Population without Selection For this example, we will consider the class to be a natural, randomly mated group with no selection and each individual being heterozygous for a particular phenotype having the genotype Aa. The initial gene frequency in this population is 0.5 for “A,” the dominant allele and 0.5 for “a,” the recessive allele. Therefore p and q both equal 0.5. Every pair of individuals should keep track of their child’s genotype after each mating. In addition, the class as a group must keep track of totals for each mating during each of the five generations. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. Work in pairs. For each generation, each pair will produce two offspring. For this example, each person in the class should obtain 4 index cards. Label two “A” and two “a.” These cards represent the only two unique gametes with a heterozygous individual with a genotype of Aa can produce. Each person in a group of two should place the index cards face down on a flat surface. Shuffle cards so that they are randomly mixed. Each person selects only one card from their pile of 4 cards and places it together with the one selected by your partner. Flip the cards over and record the genotype of this mating in Table 2. Recover your index card so again you have two of each allele “A”, and “a.” Reshuffle the cards between selections. With your partner, produce a second offspring and record the genotype. Be sure to collect the class totals on the whiteboard at the end of this generation and for the next 4 generations. Now you will assume the genotype of the first child and your partner will assume the genotype of the second child. Re-label your four index cards to reflect the gametes which you would produce with the new genotype. For example if your child was AA, your cards would be labeled A,A,A,A. Similarly, if your child was Aa, your cards would be labeled A,A,a,a. Now, things are going to get a little weird. Randomly find someone else in class to mate with. If someone approaches you with a twinkle in their eye and says, “How’s about you and me?” Your reply should be, “Absolutely.” We have to be a little promiscuous in this simulation to model real population dynamics. Random mating is key. Using the same procedure for the first generation, produce two new offspring with your new mate. Again record the genotype of each child in the table. Remember, the class as a group should also keep track of each genotype after each generation. Record the class totals. As before, one student of the pair will assume the genotype of the first child and the other student should assume the genotype of the second child produced during the mating. If necessary, re-label your index cards to reflect the four gametes which you would produce. Again, each person should randomly seek out another mate and repeat the procedure generating two new children. Record the new genotypes as before. Keep track of the class totals after each mating. This process of “random mating” without selection should continue until you have produced 5 generations of children. Be sure to place your individual results and the class totals for each mating into Table 2. Analysis 1. Calculate, from your experimental data, gene frequency values of the “A” and “a” alleles, as well as p and q in your class population. These can be obtained by using the following formulas to determine A (p) and a (q) frequencies: a) A Frequency = (Total number of AA individuals x 2) + (Total number of Aa individuals) / (Total number of all individuals) b) a Frequency = (Total number of aa individuals x 2) + (Total number of Aa individuals) / (Total number of all individuals) 2. Determine the theoretical Hardy-Weinberg frequencies of the “Class Totals for each Genotype added over 5 Generations”: 1. 2. 3. p2 (AA): __________________ 2pq (AA): _________________ q2 (aa): ___________________ 3. Determine p and q for “Class Totals for each Genotype added over 5 Generations”: 1. 2. p: ___________________ q: ___________________ 4. Determine the theoretical Hardy-Weinberg frequencies of the beginning population when p=0.5, and q=0.5. 1. 2. 3. p2 (AA): ___________________ 2pq (Aa): __________________ q2 (aa): ____________________ 5. Calculate the genotype frequencies in Part 2 from the 5th generation only: 1. 2. 3. AA: ___________________ Aa: ___________________ aa: ___________________ 6. How do the values from all of the generations added together compare to 5th generation? Have they changed? How? We’ll continue with parts 3 and 4 on Monday. Part 3: Testing an Idealized Hardy-Weinberg Population with Selection Now that you have the facts of life well in hand, we can begin to modify our simulation, making it more realistic and enable us to investigate some basic questions about selection and gene frequencies. In humans, there are several genetic “diseases” that have been thoroughly investigated. One good example is sickle-cell anemia. This is a single allele trait; the homozygous recessive genotype is lethal, and the afflicted individual usually dies prior to reaching reproductive maturity. Both the homozygous dominant and the heterozygote survive. At this point, we will consider both the homozygous dominant and the heterozygote phenotypically identical. In other words, we are selecting against the homozygote recessive 100% of the time. The procedure is similar to Part 2. Start again with your initial genotype, and choose the genotype of your offspring as in Part 2. This time, however, there is one important difference. Every time your offspring is aa it dies. Since we want to maintain a constant population size, the same two parents must try again until they produce a surviving offspring. As you might already have realized, there is one real problem that must be solved before we begin. 25% of you are already homozygous recessive or dead! There are two ways we could deal with this: (a) We could reassign genotypes so that no one starts with aa, but then our original population would not follow a Hardy-Weinberg distribution, prohibiting any comparisons between Part 2 and Part 3. (b) Instead, we will make the assumption that aa in the initial parent is not lethal, and that two aa individuals cannot mate. In other words, if you are aa, just be sure not to mate with another aa in the first round. Proceed through five generations, selecting against the homozygous recessive offspring. Now, add up the genotype frequencies that exist in the population and calculate the new p and q. Analysis 1. How do the new p and q compare to the values in Case I? 2. Has the population changed? 3. What would happen to the p and q if we went another five generations? 4. In a large population, what are the chances of eliminating completely a deleterious recessive allele? Part 4: Testing an Idealized Hardy-Weinberg Population with a Heterozygous Advantage From Part 3, it is easy to see that the lethal recessive allele rapidly decreases in the population. However, data from many human populations show an unexpectedly high frequency of the sickle-cell allele present in some populations. In other words, our simulation does not accurately reflect the real situation. The heterozygote is slightly more resistant to a deadly form of malaria than the homozygous dominant individual. In other words, there is a slight selection against the homozygous dominant individual as compared to the heterozygote. This fact is easily incorporated into our simulation. In this round, keep everything the same as in Part 3, except that if your offspring is AA, flip a coin. Heads he lives, tails he dies of malaria, and the same parents must mate again until they get a viable offspring. Go through five generations, starting again with your initial genotype. The genotype aa dies 100% of the time. After five generations, total the genotypes and calculate p and q. Now, starting with the genotype you last had, go through five more generations, and again total the genotypes and calculate p and q. If time permits, another five generations is extremely informative. Analysis 1. How do p and q in Part 3 compare with Part 2 and Part 4? 2. Do you think the recessive allele will be completely eliminated in either Part 3 or Part 4? 3. What is the importance of heterozygous advantage in maintaining genetic variation in populations? Part 5 (Time permitting) – Genetic Drift In Part 2, the role of small populations in genetic drift was suggested. It is possible to use our simulation to look at this phenomenon in more detail. Divide the lab into several smaller populations (For example, a class of 30 could be divided into 3 populations of 10 each.) When the populations are reproductively isolated by mountain ranges, oceans, or imaginary walls, total the genotypic and allelic frequencies in each population. Now go through five generations as in Part 2. Record the new genotypic frequencies, and calculate the new p and q for each population. Analysis 1. How did the initial genotypic frequencies of the populations compare? 2. What do your results indicate about the importance of populations size as an evolutionary force?