Date of Sowing and Nursery Growth of Provenances of Pseudotsuga... Fertilizer Regimes
advertisement

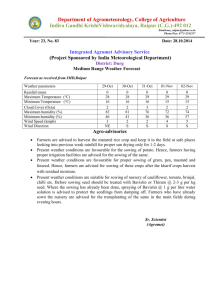
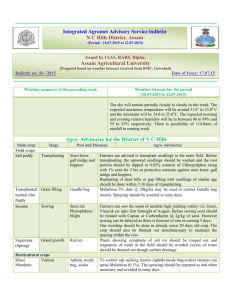
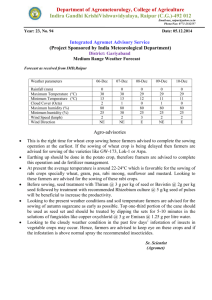
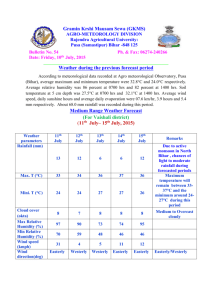
Date of Sowing and Nursery Growth of Provenances of Pseudotsuga menziesii Given Two Fertilizer Regimes Author(s): Frank C. Sorensen Source: Journal of Applied Ecology, Vol. 15, No. 1 (Apr., 1978), pp. 273-279 Published by: British Ecological Society Stable URL: http://www.jstor.org/stable/2402936 . Accessed: 20/01/2015 19:49 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact support@jstor.org. . British Ecological Society is collaborating with JSTOR to digitize, preserve and extend access to Journal of Applied Ecology. http://www.jstor.org This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions Journal of Applied Ecology (1978), 15, 273- 279 DATE OF SOWING AND NURSERY GROWTH OF PROVENANCES OF PSEUDOTSUGA MENZIESII GIVEN TWO FERTILIZER REGIMES BY FRANK C. SORENSEN Forestry Sciences Laboratory, Pacific Northwest Forest and Range Experiment Station United States Department of Agriculture, Corvallis, Oregon, U.S.A. SUMMARY (I) Seed was collected from natural stands of Pseudotsuga menziesii growing at 250, 520 and 880 m elevation in Oregon, U.S.A. Seed was also obtained from a cross between trees at 250 and 520 m. The seed was sown on five dates between 23 April and 8 July 1974. Half the plots were left unfertilized in 1974; all plots were fertilized in 1975. (2) Early sowing significantly increased stem height and diameter, advanced the date of budset and altered the period of stem elongation, extending it in the fir st year and shortening it in the second. (3) Fertilizer increased seedling height and diameter and the elongation period if seeds were sown early but not if they were sown late. (4) There were significant differences among provenances from different elevations for stem diameter, date of budset and date of budftush. (5) Interactions between provenances and dates of sowing were significant for dates of budset and budftush, and for stem height, diameter and form. Comparisons of young material of different provenances must take into account the effect of cultural conditions, particularly conditions altering the length of the elongation period. INTRODUCTION Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) is native to the north temperate region. Its seed germinates in the spring at a time when daylength and soil and air temperature are increasing and precipitation, humidity and cloud cover are decreasing (Lavender 1958). Presumably the pattern of changing response to environmental factors during germination and establishment is adapted to the pattern of changing environmen­ tal conditions during spring and early summer. If this is true, then altering the date of sowing could have important effects on seedling development. Many workers have investigated the effects of sowing date on a wide range of plant traits in a number of crops and weeds but few studies have been made on trees. Baak (1933) and Baak & Hilf (1932) studied the effect of autumn and spring sowing on Douglas-fir. After two growing seasons, autumn-sown plants were 21% taller and over twice as heavy (Baak 1933). Similar results were obtained by Demeritt & Hocker (1975) working with eastern white pine. Differences of I week in the time of germination affected top dry weight after I year in some provenances. Weather and organizational problems have unavoidably influenced the date of sowing in outdoor nurseries. More recently, new technologies in bed house culture have added the possibility of deliberately altering sowing dates. This paper reports information on the 0021-8901(78/0400-0273$02.00 ©1978 Blackwell Scientific Publications 273 This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions 274 Growth of provenances of Douglas-fir effect of sowing date on Douglas-fir growth and development. The growth and developmental timing of seedlings of different provenances have been investigated, and the effect of fertilizer application on those responses has been studied. Stem elongation has been examined, with special reference to the length of the elongation period and the time when elongation occurs. Observations made during the second growing season have been analyzed, _and the importance of 'carryover' effects considered. MATERIALS AND METHODS Source of material The seed came from trees growing at three elevations (250, 520 and 880 m) in the Oregon Cascade Range at 45°N, and from crosses between trees at two locations (pollen from trees at 250m applied to flowers on trees at 520 m). Each location sample (hereafter called provenance) was represented by eight progenies, either derived from crosses among trees-within-location (250, 520 and 880 m), or from single crosses between trees at the two lower elevations. Equal numbers of pre-germinated seeds from each family were counted out and bulked before sowing; the identity of individual families was not maintained in the test. Treatments The seeds were pregerminated in the laboratory to a radicle length of0·5- 2·0 em then sown into a raised (0· 5 m) coldframe outdoors at 19-day intervals between 23 April and 8 July 1974. Fertilizer was applied to half the plots in 1974 and to all plots in 197 5. A slow-release fertilizer (14-7-7) was applied at a rate of 56 kg N ha- 1 , l week before plan tin g. The fertilizer was placed about 15 em below the soil surface of fertilized plots, then all plots were watered heavily. Additional applications of liquid fertilizer (20-19-18) were given to fertilized plots at an equivalent rate of N at each subsequent sowing, and at 21-day intervals after the last sowing until early September. In 1975, liquid fertilizer (23-19-17) was applied to all plots every 21_days from early April until mid-August. The coldframes were shaded (50% sunlight) from sowing until mid-September in 1974 and from late May until early September 1975. Water was applied three to four times daily immediately after sowing, decreasing to one heavy application daily by the end of summer 1974. In 1975, beds were watered heavily every l-4 days. Measurements The following measurements were made in 1974: (a) number of days from date of sowing until seedcoat dropped from cotyledons (observed daily); (b) budset, i.e. the date on which a developing terminal bud could first be seen on the main stem (observed weekly); (c) elongation period (days between date of sowing and date of budset); (d) epicotyllength (elongation of the main stem during the first year); (e) elongation rate (epicotyllength/elongation period). The following measurements were made in 1975: (a) budftush (date on which green needles could first be observed); (b) budset (see measurement (b) for 1974); This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions FRANK C. SORENSEN 275 (c) Elogation period (number of days between budftush and budset); (d) 2-year height (total elongation of the mainstem during the 2 years); (e) elongation rate (second-year elongation/second-year elongation period); (f) diameter (stem diameter below the cotyledons); (g) stem form (height/diameter). Experimental design and analysis A double split-plot design with three replications was used. The main plots were fertilized v. nonfertilized, surrounded by border rows and separated by 30 em of bare ground. Provenances were assigned at random to sub-plots, and dates of sowing assigned at random to sub-sub-plots. Spacing between seedlings was 7·6 em. The analysis of variance tested for linear and nonlinear trends associated with date of sowing alone and in interaction with fertilizer and provenance. The curves in the figures are best fits to polynomials using a least squares procedure. RESULTS First-year measurements Earlier sowing caused slower initial development of the seedlings. Plants from earlier sowings took longer to shed seed coats; the linear regression of days to shedding on date of sowing accounted for 89% of total variance. The average number of days to shed seedcoats was closely correlated with the average daily air temperature (r=0·99). The date ofbudset was earlier and the elongation period longer when seeds were sown early (Fig. l(a)). Fertilizer delayed budset and lengthened extension period if seeds were sown early, but had the opposite effect if seeds were sown late (Fig. I (a)); the fertilizer x date of sowing interaction was significant (P< 0·01). Epicotyl lengths at the end of the first growing season were greater for plants sown early; the linear regression accounted for 43% of the variation. Fertilizer increased epicotyllength if seeds were sown early, but had negligible effect if they were sown late (Fig. l(b)) (date-of-sowing x fertilizer interaction, P<O·OOJ). First-year height was reduced about 1·9 and 1·1 mm, on fertilized and non-fertilized plots respectively, for each day delay in sowing between 23 April and 12 May, and by about 0·5 and 0·1 mm respectively, for each day delay in sowing between 12 May and 19 June. Second-year measurements Date of budset in the second year was 17 days earlier (P < 0·00 1) for plants that had been sown in April of the previous year than when sown in July; the elongation period was 18 days shorter (P<O·OOl); linear regressions accounted for 32% and 35% of the variation, respectively. Seedlings from seeds collected at 250m flushed 5 days earlier than those from 880 m (P < 0·001). The budset dates of the various provenances were affected differently by date of sowing (Fig. 2(a)) (date-of-sowing x provenance interaction, P < 0·01 ). Seedlings from 520 m had about the same developmental timing regardless of sowing date; seedlings from the other elevations did not. Early-sown plants were taller, larger in diameter and had a smaller height/diameter ratio (P < 0·001) after 2 years, linear regression accounting for 43%, 74% and 34% of the variation, respectively. This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions 276 Growth of provenances of Douglas-fir 23 April Dote of sow1ng FIG . I. Effect of date of sowing on date of first-year budset (a) and leader extension (b) of Douglas-fir seedlings receiving (___.) or not receiving (o -- o) fertilizer. Curves are polynomials fitted by the least squares procedure. S.E. (S) is the standard error for comparing sowing·dates and S.E. (F) for fertilizer treatments. Fertilizer application increased diameter and decreased height/diameter ratio of plants sown early, but not of those sown late (date-of-sowing x fertilizer interaction, P< 0·001). Each day delay in date of sowing decreased total height by an average of 1·6 mm and diameter by 0·023 mm (fertilized plots) or 0·018 mm (non-fertilized plots). Provenances did not differ in average second-year height, but they did differ in response to date of sowing (Fig. 2(b)) (date-of-sowing x provenance interaction, P< 0·01). For example, seedlings from 880 m were taller than those from 520 m (45·1 v. 38·8 em) when sown 23 April, but shorter (27·7 v. 30·7 em) when sown 8 July. Second-year diameter of the provenances showed a similar interaction with date of sowing (P< 0·01 ). DISCUSSION Early sowing increased plant size. The amount of increase was affected by the length of the elongation period, the time when elongation occurred, fertilizer application and provenance. This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions FRANK 277 C. SORENSEN (a} Aug . 30 ----- -n Aug 25 ~ Aug 20 "0 ..5 Aug 15 ~ Aug . 10 8 Aug.5 July 30 30 rs.E. IS.E. (S) (P) Date of sowmg 2. Effect of date of sowing on date of second-year budset (a) and plant height (b) of Douglas-fir seedlings from 250m(---), 520m (o--o), 880 m ("--")and the cross 250 x 520 m (o--- -o). The curves are polynomials fitted by the least square procedure. S.E. (S) is the standard error for comparing sowing dates and S.E. (P) for comparing provenances. FIG . Elongation period First year Plants grow exponentially as long as new growth includes productive photosynthetic capital. In first-year seedlings of Douglas-fir, photosynthetic capital is produced primarily when the leader is elongating. In this test, plants from the 23 April sowing had a 15% longer elongation period than plants sown 12 May, and their epicotyllength was 49% greater. However, if plants from later sowings are compared, the benefits of a longer growing season are not so pronounced. Plants sown 31 May had a I 0% longer elongation period than those sown 19 June and their epicotyllength was II% greater. Thus, the time when elongation occurred, as well as the length of the elongation period, influenced plant height. Studies on the photosynthetic characteristics of Douglas-fir seedlings suggest reasons This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions 278 Growth of provenances of Douglas-fir for the advantages of early spring sowing. The temperature optimum for photosynthesis is relatively low, when seedlings are in the cotyledon stage, and increases with age in the first few weeks of seedling development (Sorensen & Ferrell 1973). For seedlings about 5 weeks of age and older, temperature and moisture appear to be more suitable for photosynthesis and growth in late spring than in summer (Brix 1967; Krueger & Ferrell 1965; Zavitkovski & Ferrell 1968). Leaf production in mid-summer may even be slowed down by high temperatures (Sorensen & Ferrell 1973). Growth should be greatest if leaf production and leaf area are as large as possible when climatic conditions are also more favorable for photosynthesis (Watson 1956). Early sowing apparently matched the climatic requirements of the seedlings to the climate of the area better than did later sowing. Second year The effects of early sowing on seedling size continued through the second year, presumably because of differences in photosynthetic capital which were present during bud dormancy and at the beginning of the second growing season. This greater photosynthetic capital produced greater elongation. For example, plants from the 23 April sowing elongated 9% more than those from the 12 May sowing, even though the elongation period of the former was 10 days less. Diameter growth and plant form appeared to be regulated primarily through an effect of sowing-date on date of budset. Much diameter increment occurs after budset and, in our nursery, has been measurable in all months except January (unpublished data). Early budset allowed more time for diameter growth, and the time added was in late summer and early autumn when conditions were favorable for assimilation. Because early sowing caused early budset in both growing seasons, it resulted in plants with smaller height-diameter ratios even though the plants were tall. Fertilizer Fertilizer application increased seedling height if plants were sown early, but not if they were sown late (Fig. l(b)). The increase in growth was probably due, at least partly, to an effect of fertilizer on date of budset, for fertilizer delayed budset of early-sown but not late-sown plants (Fig. l(a)). Except for second-year diameter, fertilizer application did not affect differences among provenances. This test and others (Campbell & Wilson 1973; Campbell & Sorensen, in preparation) indicate that provenances may interact more with treatments which change the timing of growth, e.g. different dates of sowing or different temperature environ­ ments, than with treatments which generally do not change the timing of growth, e.g. fertilizer or spacing. Provenance The responses of provenances to date of sowing were not interpretable simply in terms of the elevation of their site of origin. On the basis of height and diameter measurements, seedlings from 880 m were most sensitive to sowing date, seedlings from 520 m least. On the basis of date of budset, seedlings from 520 m were again least sensitive to date of sowing, but seedlings from -the elevational cross were the most sensitive. The natural elevational range of Douglas-fir, in the region where the seeds were collected, spans about 1400 m. The sample oflocations in the test covered only about 600 m; This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions FRANK C. SoRENSEN 279 all were in an elevational band with a moderately long growing season. Had a greater elevational range been sampled, perhaps the pattern of response would have been more easily related to the elevation of the site of origin. It seems possible that provenances from low elevations, which have a long growing season, would be more sensitive to date of sowing than those from high elevations, which usually have a short elongation period. CONCLUSION (1) Because of interactions between provenances and sowing dates, comparisons between provenances, using young material, should be sown on more than one date, preferably spanning the range of sowing dates used in practice. (2) Date of budset, as well as affecting susceptibility to frost, influences elongation period and, in turn, seedling size. Cultural treatments which interact with provenances in their effect on budset date probably also will interact in their effect on seedling size. Therefore, first-year budset and elongation period observations should be good early indicators of provenance cultural-treatment interactions in growth. ACKNOWLEDGMENT The author appreciates the thoughtful and helpful comments of the two referees. REFERENCES Baak, W. (1933). Die Nachzucht der Douglasie. Forstarchiv, 9, 106. Baak, W. & Hilf, H. H. (1932). Die Nachzucht der Douglasie. Forstarchiv, 8, 289- 91. Brix, H. (1967). An analysis of dry matter production of Douglas-fir seedlings in relation to temperature and light intensity. Canadian Journal of Botany, 45, 2063-72. Campllell, R. K. & Wilson, B. C. (1973). Spacing-genotype interaction in Douglas-fir. Si/vae Genetica, 22, 15- 20. Demeritt, M. E., Jr & Hocker, H. W., Jr. (1975). Influence of seed weight on early development of eastern white pine. Northeastern For. Tree Improvement Conference Proceedings, 22, 130-7. Krueger, K. W. & Ferrell, W. K. (1965). Comparative photosynthetic and respiratory responses to temperature and light by Pseudotsuga menziesii var. menziesii and var. glauca seedlings. Ecology, 46, 794-801. LaveDIIer, D.P. (1958). Effect of field stratification on Douglas-fir seed germination. Research Note, 33, 8 pp. Oregon For. Lands Res. Cent., Corvallis . Sorensen, F. C. & Ferrell, W. K. (1973). Photosynthesis and growth of Douglas-fir seedlings when grown in different environments. Canadian Journal of Botany, 51, 1689- 98. Watson, D. J. (1956). Leaf growth in relationship to crop growth. The Growth of Leaves (Ed. by F . L. Milthorpe), pp. 178-91. Butterworth's Scientific Publications, London. Zavitkovski, J. & Ferrell, W. K. (1968). Effect of drought upon rates of photosynthesis, respiration, and transpiration of seedlings of two ecotypes of Douglas-fir. Botanical Gaiette, 129, 346-50. (Received 29 June 1977) This content downloaded from 166.6.105.57 on Tue, 20 Jan 2015 19:49:30 PM All use subject to JSTOR Terms and Conditions