Electronic Supplementary Information for Roffler et al. “Lack of sex
advertisement

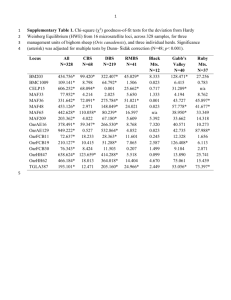
Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Table 1. Primer name, tail, and primer sequence for the 15 nuclear microsatellite loci and 3 mitochondrial DNA genes used on Dall’s sheep from Wrangell-St. Elias National Park and Preserve, Alaska. Primer name Locus or Locus Type1 Gene Tail2 Primer Sequence (5’ – 3’) Source Neutral nuclear loci MAF36 µsat Dinucleotide repeat polymorphism Direct F: TATACCTGGGAGGAATGC R: AGTTGGACACAATTGAGC Swarbrick et al. 1991 AE16 µsat Dinucleotide repeat polymorphism Direct F: GAGGAATGGGTGAAGACG R: ATGGCTCGGTAATATTCC Penty et al. 1993 HH62 µsat Dinucleotide repeat polymorphism Direct F: AGAGAAAAGCTGGGGTGC R: CGAGTCAAACACTACTGAG Ede et al. 1994 MAF209 µsat Dinucleotide repeat polymorphism Direct F: GCACTTAAGTATGTAGGATGC R: AGTTGGATACAACCGTGG Buchanan and Crawford 1992 MAF33 µsat Dinucleotide repeat polymorphism Direct F: CAGAGAAAGATACACGTGC R: CTCACTCTTTCTGTCTCTG Buchanan and Crawford 1992 MAF48 µsat Dinucleotide repeat polymorphism Direct F: GACTGAGCAACTAAGTACG R: AAGCCACTTTTCAGATGC Buchanan et al. 1991 FCB266 µsat Dinucleotide repeat polymorphism Direct F: ACCAAACACACAGCCTGC R: CCACTAGCTTTACATAGG Buchanan and Crawford 1993 FCB304 µsat Dinucleotide repeat polymorphism Direct F: TGCTGTCAACTGGGTCAG R: GGAGCTTTCAATAAAGAATCGG Buchanan and Crawford 1993 1 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Table 1, cont. Primer name, tail, and primer sequence for the 15 nuclear microsatellite loci and 3 mitochondrial DNA genes used on Dall’s sheep from WrangellSt. Elias National Park and Preserve, Alaska. Loci linked to nuclear functional genes3 KERA µsat Corneal keratin sulfate proteoglycan Direct + M13F(-29) F: GTACTGAACCAAATAGTACAGCAG CCAAT R: GCATGGCAACCCACTCCAGTAT Tasheva et al. 1998 KRT2 µsat Intermediate filament type II keratin Direct + SP6 F: GCCTCTAGGCGTGAGGGTTTT R: AAGGGCCAAGAGTCATTCACAT Powell et al. 1993 SOMAb µsat Grunniens growth hormone receptor Direct F: ATGTTCTAATCTTATCTGGTACCAGG R: GTCCTCCCCAAATCAATTACATTTTCTC Ma et al. 2010 ADCYAP1 µsat Adenylate cyclaseactivating polypeptide 1 Direct F: CCAGACGCCGACTTCGCCGAGG R: GCCTGAAGTCCACTGAGAAGAAAGGA Moore et al. 2006 MMP9 µsat Matrix Metallo Proteinase 9 Direct F: CTTGCCTTCTCATGCTGGGACT R: GTGAGGATAGCAACTCTTGGTCTGGCT Adamson et al. 2000 OLADRBps µsat DRB = MHC class II B MHCII(F)SP6 MHCII(R)4 MHCII(R) F: CTGCCAATGCAGAGACACAAGA R4: TGTCTTGTCATCTCTACG R: GTCTGTCTCCTGTCTTGTCATC Scott et al. 1992 TCRBV62 µsat T-cell receptor Direct F: TGAGTCCTCAGCAAGCAGGT R: ACTGGGACACTACTCCAGCTCTT Buitkamp et al. (unpublished) 2 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Table 1, cont. Primer name, tail, and primer sequence for the 15 nuclear microsatellite loci and 3 mitochondrial DNA genes used on Dall’s sheep from WrangellSt. Elias National Park and Preserve, Alaska. Loci in mitochondrial DNA COX1F COX1R mtDNA sequence Cytochrome oxidase I M13F(-29) M13R F: CTGTTCTTAGATTTACAGTCTA R: CTGCCAGATGTAGGGAGAAAAT This study: designed from GenBank Accession EF490456 (O. aries) CYTB_F CYTB_R mtDNA sequence Cytochrome b M13F(-29) M13R F: CCCCACAAAACCTATCACAAA R: AGGGAGGTTGGTTGTTCTCC Pedrosa et al. 2005 CYTB_IN_F CYTB_IN_R mtDNA sequence Cytochrome b M13F(-29) M13R F: ACCTCCTTTCAGCAATTCCA R: CCTGTTTCGTGGAGGAAGAG Pedrosa et al. 2005 OdaProL OvisCR4H OvisCR5H mtDNA sequence Control region M13F(-29) M13R M13R F: ACTATCAACACCCAAAGC R: CTAGTGGACAGGATACGC R: GGATTTGACTTTATGTGC This study; designed from GenBank Accession AY858379, FJ545782 (O. aries) and DQ249916 (O. nivicola) 1 µsat = microsatellite locus; mtDNA = mitochondrial DNA 2 forward primer of each microsatellite pair, and both primers for mtDNA sequencing were synthesized with an additional 19-20 nt M13F, M13R, or SP6 universal tail on the 5’ end (M13F [-29]: CACGACGTTGTAAAACGAC; SP6:GATTTAGGTGACACTATAG; M13R: GGATAACAATTTCACACAGG) 3 4 Luikart et al. 2008 Redesigned for this study. The redesign reverse is 10 nt smaller than original 3 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Primer references Adamson R, Logan M, Kinnaird J, Langsley G, Hall R (2000) Loss of matrix metalloproteinase 9 activity in Theileria annulata-attenuated cells is at the transcriptional level and is associated with differentially expressed AP-1 species. Molecular Biochemical Parasitology, 106, 51–61. Buchanan FC, Swarbrick PA, Crawford AM (1991) Ovine dinucleotide repeat polymorphism repeat polymorphism at the MAF48 locus. Animal Genetics, 22, 379–380. Buchanan FC, Crawford AM (1992) Ovine microsatellite polymorphism at the MAF70 locus. Animal Genetics, 23, 185. Buchanan FC, Crawford AM (1993) Ovine microsatellites at the OarFCB11, OarFCB128, OarFCB193, OarFCB266 and OarFCB304 loci. Animal Genetics, 24, 145. Buitkamp J, Schwaiger F-W, Epplen J-T. Vb6 T-cell receptor elements in artiodactyls; Conservation and germline polymorphisms. Unpublished. Ede AJ, Peirson CA, Henry H, Crawford AM (1994) Ovine microsatellites at the OarAE64, OarHH22, OarHH56, OarHH62, and OarVH4 loci. Animal Genetics, 25, 51–52. Luikart G, Pilgrim K, Visty J, Ezenwa VO, Schwartz MK (2008) Candidate gene microsatellite variation is associated with parasitism in wild bighorn sheep. Biology letters, 4, 228–31. Ma Z, Xu J, Zhong J, Dou Q, et al. (2010) Structural features of the 5’ flanking region of the Yak (Bos grunniens) growth hormone receptor (GHR) gene. Archiv Tierzucht, 53, 372–378. Moore S, Alexander L, Brownstein M, Guan L. et al. (2006) BC Cancer Agency, Canada’s Michael Smith Genome Sciences Centre, Vancouver, British Columbia, Canada. Pedrosa S, Uzun M, Arranz J et al. (2005) Evidence of three maternal lineages in near eastern sheep supporting multiple domestication events. Proceedings Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 272, 2211–2217. Penty JM, Henry HM, Ede AJ, Crawford A (1993) Ovine micro- satellites at the OarAE16, OarAE54, OarAE57, OarAE119 and OarAE129 loci. Animal Genetics, 24, 219. Powell BC, Crocker LA, Rogers GE (1993) Complete sequence of a hair-like intermediate filament type II keratin gene. Mitochondrial DNA, 3, 401–405. 4 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Scott PC, Maddox JF, Gogolin-Ewens KJ, Brandon MR (1992) The nucleotide sequence and evolution of ovine MHC class II B genes: DQB and DRB. Immunogenetics, 35, 217. Swarbrick PA, Buchanan FC, Crawford AM (1991) Ovine dinucleotide repeat polymorphism at the MAF36 locus. Animal Genetics, 22, 377–378. Tasheva ES, Funderburgh JL, Corpuz LM, Conrad GW (1998) Cloning, characterization and tissue-specific expression of the gene encoding bovine keratocan, a corneal keratin sulfate proteoglycan. Gene, 218, 63– 68. 5 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Table 2. Locus-specific genetic variation at 15 microsatellite markers genotyped on Dall’s sheep including the total number of alleles observed (A), mean observed (Ho) and expected heterozygosity (He) across the study area, Wrangell-St. Elias National Park and Preserve, Alaska. Bold values are significant at the 0.05 level for deviation from Hardy-Weinberg proportions. CHIS ECHU NUTZ NWR STE SWR WCHU A Ho He A Ho He A Ho He A Ho He A Ho He A Ho He A Ho He MAF36 5 0.750 0.667 4 0.750 0.561 4 0.609 0.658 6 0.667 0.717 6 0.531 0.553 5 0.661 0.703 3 0.500 0.390 AE16 3 0.625 0.650 3 0.259 0.230 3 0.652 0.655 3 0.435 0.480 4 0.422 0.536 3 0.471 0.577 3 0.333 0.287 HH62 6 0.720 0.729 4 0.429 0.363 6 0.667 0.756 7 0.688 0.686 7 0.541 0.695 6 0.821 0.764 4 0.750 0.666 MAF209 2 0.240 0.269 4 0.357 0.391 3 0.478 0.481 5 0.440 0.466 6 0.535 0.594 3 0.273 0.301 3 0.563 0.506 MAF33 4 0.636 0.538 3 0.464 0.541 3 0.476 0.373 5 0.500 0.540 3 0.552 0.663 4 0.569 0.546 4 0.733 0.727 FCB266 8 0.667 0.733 4 0.556 0.497 5 0.609 0.641 7 0.735 0.724 5 0.430 0.571 5 0.429 0.439 4 0.533 0.496 KRT2 5 0.577 0.669 3 0.630 0.549 4 0.609 0.710 5 0.490 0.537 5 0.539 0.583 4 0.321 0.310 3 0.267 0.238 KERA 3 0.154 0.144 1 0.000 0.000 1 0.000 0.000 2 0.180 0.164 1 0.000 0.000 2 0.018 0.018 1 0.000 0.000 SOMAb 10 0.750 0.779 7 0.615 0.746 7 0.591 0.764 12 0.744 0.772 9 0.511 0.554 8 0.760 0.767 4 0.688 0.635 ADCYAP1 6 0.731 0.761 3 0.679 0.563 7 0.857 0.786 7 0.680 0.741 5 0.620 0.630 6 0.574 0.502 4 0.667 0.629 MMP9 5 0.680 0.609 5 0.741 0.731 6 0.609 0.741 7 0.727 0.792 8 0.787 0.805 7 0.709 0.766 6 0.800 0.704 MAF48 5 0.500 0.521 4 0.593 0.676 4 0.696 0.589 5 0.720 0.635 5 0.600 0.646 5 0.518 0.598 3 0.625 0.623 FCB304 4 0.538 0.575 2 0.464 0.499 3 0.435 0.506 3 0.540 0.521 5 0.430 0.540 4 0.571 0.560 3 0.875 0.635 OLADRBps 8 0.808 0.818 5 0.333 0.408 8 0.826 0.803 8 0.592 0.750 7 0.394 0.524 5 0.596 0.672 4 0.625 0.602 TCRBV62 3 0.440 0.394 2 0.214 0.245 3 0.318 0.491 4 0.540 0.543 3 0.333 0.430 3 0.482 0.490 3 0.500 0.525 6 Electronic Supplementary Information for Roffler et al. “Lack of sex-biased dispersal promotes fine-scale genetic structure in alpine ungulates”. Figure 1. Autocorrelogram of the spatial coefficient, r, as a function of geographic distance for Dalls’ sheep (males, grey dashed line; females, black dashed line; and both sexes combined, black solid line), Wrangell-St. Elias National Park and Preserve, Alaska, 2007-2009. Significant spatial genetic autocorrelation can be assumed when mean r exceeds the permuted 95% confidence intervals (dashed lines) and the bootstrapped 95% confidence error bars for each distance class do not intercept the X-axis of r = 0. 7