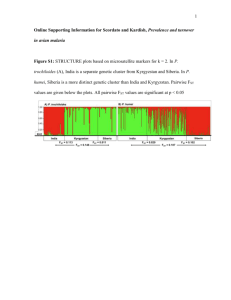
Online Resource 1 Eighteen microsatellite markers were assayed
advertisement

Online Resource 1 Eighteen microsatellite markers were assayed initially using primers developed for the roseate tern (Sterna dougallii; Szczys et al. 2005), red-billed gull (Larus novaehollandiae; Given et al. 2002), and black-legged kittiwake (Rissa tridactyla; Tirard et al. 2002). These primers have been used successfully in population genetic studies of other tern species including the South American tern (Sterna hirundinacea; Faria et al. 2010) and the common tern (S. hirundo; Sruoga et al. 2006). Initial tests of these loci in a subset of Caspian terns failed to yield reliable products for seven loci (K6, K56, K67, K71, RBG20, RBG28, RBG39) and five other loci were not polymorphic (K16, K31, K32, RBG13, Sdaat27). The six remaining loci were PCR amplified either individually (Scaac20, RBG18) or in di-locus multiplexes (RBG27/RBG29 and Sdaat20/Sdaat46). The reaction conditions for most PCRs were identical to those outlined in Taylor et al. (2010). For locus Scaac20, only 0.06 μM WellRED D4 labelled M13 was used and the number of PCR cycles was increased by ten. Allele sizes were determined on a BeckmanCoulter (Mississauga, ON) CEQ 8000 genetic analysis system following the protocols of Sun et al. (2009). Due to difficulties interpreting the stutter pattern of locus Sdaat46, it was excluded from all further analysis. Approximately 490 base pairs of the mitochondrial cytochrome b gene was amplified from each individual using general avian primers b5 (5'TTCCACCCCTACTTCTCACTAAAAGA-3'; T. Birt, unpublished data) and b6 (5'GTCTTCAGTTTTTGGTTTACAAGAC-3'; T. Birt, unpublished data). PCRs contained approximately 5 ng of DNA, 1 x Multiplex Mix (Qiagen, Mississauga, Ontario), and 0.7 mM of each primer under standard conditions with annealing at 50 °C for 30 seconds. PCR products were sequenced with both primers on a 3730XL DNA Analyzer (Applied Biosystems, Foster City, California) at the Genome Quebec Innovation Centre (McGill University, Montreal, Quebec). Chromatograms were verified manually and sequences were aligned using ClustalW (Thompson et al. 1994) as implemented in BioEdit (Version 7.0.5.3; Hall 1999). Literature Cited Given AD, Mills JA, Baker AJ (2002) Isolation of polymorphic microsatellite loci from the redbilled gull (Larus novaehollandiae scopulinus) and amplification in related species. Mol Ecol Notes 2:416-418 Faria PJ, Campos FP, Branco JO, Musso CM, Morgante JS, Bruford MW (2010) Population structure in the South American tern Sterna hirundinacea in the South Atlantic: two populations with distinct breeding phenologies. J Avian Biol 41:378-387 Sruoga A, Butkauskas D, Prakas P, Paulauskas A (2006) Evaluation of the genetic structure of the breeding common tern (Sterna hirundo) population by means of microsatellite markers. Biologija 1:47-52 Sun Z, Gomez-Diaz E, Bailie A, Friesen V (2009) Isolation and characterization of microsatellite loci for storm-petrels. Mol Ecol Res 9: 913-915 Taylor SA, Morris-Pocock JA, Sun Z, Friesen VL (2010) Isolation and characterization of ten microsatellite loci in Blue-footed (Sula nebouxii) and Peruvian Boobies (Sula variegata). J Ornithol 151:525-528 Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties, and weight matrix choice. Nucleic Acids Res 22: 4673-80 Tirard C, Helfenstein F, Danchin E (2002) Polymorphic microsatellites in the black-legged kittiwake Rissa tridactyla. Mol Ecol Notes 2:431-433