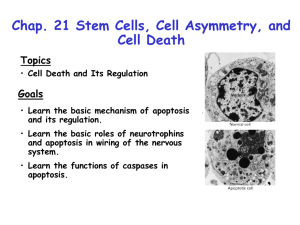
Apoptosis
advertisement

APOPTOSIS Apo: Greek: from, away Ptosis: Greek: fall, falling A specific type of cell death first described in 1972 by Kerr, et al in the British Journal of Cancer Programmed Cell Death (Lockshin and Williams, 1964) Developmental Event, may or may not be apoptotic In all our tissues, the size of individual cell populations is governed by cell birth brought about by division of cells - mitosis - and by a controlled cell deletion or 'suicide' process known as apoptosis. The name apoptosis is derived from the ancient Greek term meaning 'the falling of leaves from trees or petals from flowers'. During embryonic development, we are fashioned not only through mitosis, cell differentiation and cell migration, but also through the 'useful cell death' that is apoptosis. Numerous tissues within our adult bodies are maintained by a fine balance between mitosis and apoptosis. For example, immune responses to infection are boosted by cell division and finely tuned and curtailed by apoptosis; cells of the gut and skin are renewed through cycles of mitosis and apoptosis; the lactating breast regresses post-lactation by apoptosis. Apoptotic B Cells •Essential for proper development of the multicellular organism •"Webbed" tissue between the digits of developing human embryos is removed by apoptosis •Tadpole tail removed by apoptosis during development •Neuronal cell death by apoptosis is fundamental for CNS development • •Essential for the proper functioning of the mature organism • • • Cells of the intestinal wall die by apoptosis to be replaced by new cells Skin cells (keratinocytes) undergo apoptosis and migrate to the surface where they form the protective outer layer of skin •Essential for the removal of cells that threaten homeostasis •Virus-infected cells .Cells whose DNA has been damaged by UV light, exposure to radiation, chemotherapy •Autoreactive T cells with the potential to attack "self" are removed by apoptosis •At the termination of an immune response when they are no longer needed Apoptosis or Programmed Cell Death (PCD) A genetically controlled process for cells to commit suicide. Distinct morphological and biochemical signatures of apoptosis are: DNA fragmentation Chromatin condensation Cell shrinkage Plasma membrane blebbing. The term ‘apoptosis’ was coined by Kerr, Wyllie and Currie (1972): to distinguish between the cell deaths that occur in homeostasis and pathological cell deaths such as Trauma and ischemia. Necrosis occurs when cells are exposed to extreme physiological Conditions result in damage to the plasma membrane. Usually triggered by agents like complement and lytic viruses, hypothermia, hypoxia Necrosis Apoptosis Morphological changes • Loss of membrane integrity. • Begins with swelling of cytoplasm and mitochondria. • No vesicle formation, complete lysis • Membrane blebbing, but no loss of integrity. Aggregation of chromatin at nuclear memb. • Begins with shrinking of cytoplasm and condensation of nucleus. • Formation of membrane bound vesicles (apoptotic bodies) Biochemical changes • No energy requirement passive process • Random digestion of DNA • Postlytic DNA fragmentation • Energy (ATP)-dependent • Non-random mono- and oligonucleosomal length fragmentation of DNA. Laddering pattern • Prelytic DNA fragmentation Physiological changes • Phagocytosis by macrophages • Phagocytosis by adjacent cells or macrophages • Significant inflammatory response • No inflammatory response Apoptosis in Development and Disease : Role in animal development: 1. Sculpting structures e.g. formation of digits, neural tubes. 2. Deleting unneeded structures e.g. vestigial structures like pronephric tubes 3. Controlling cell numbers like neurons and oligodendrocytes. 4. Function as quality control, eliminating harmful cells e.g self reacting lymphocytes 5. Producing differentiated cells without organelles. E.g formation of RBC, differentiated keratinocytes. Role in Diseases: • Over-active apoptosis: examples are neurodegenerative diseases (eg Alzheimer's), immunodeficiency diseases (eg. AIDS), Stroke and coronary heart disease. • Under-active apoptosis: examples are cancer, auto-immune and chronic inflammatory diseases (eg rheumatatoid arthritis). The latter may be caused by defects in the mechanisms of apoptotic-cell clearance. Apoptosis is an Essential Process • Apoptosis (programmed cell death) plays an important role in normal development and homeostasis • Apoptosis is activated through two principal signaling pathways: intrinsic and extrinsic • Cancer is often initiated by DNA damage • Normal cells undergo apoptosis in response to stress-inducing events in the cell, such as DNA damage • Dysregulation of apoptosis is critical for cancer development and tumor cell survival Cell Death Pathways The Nobel Prize in Physiology or Medicine 2002 Sydney Brenner of the Salk Institute used the nematode Caenorhabditis elegans, which became a multicellular model experimental system, to follow cell division and differentiation from the fertilized egg to the adult via microscopic observation. He demonstrated that a specific gene mutation, induced by ethyl methane sulfonate, could be linked to a specific effect in nematode organ development. His work on nematodes created an experimental system that laid the foundation for the study of apoptosis. John Sulston of the Wellcome Trust Institute in England mapped cell lineages, where every cell division and differentiation could be followed in the development of C. elegans. There are only 959 cells in an adult nematode. He showed that specific cells lineages (nerves) undergo programmed cell death, as an integral part of the normal differentiation process. Robert Horvitz of MIT discovered and characterized key genes controlling cell death in C. elegans. He identified the first two bona fide "death genes", ced-3 and ced-4. Functional ced-3 & ced-4 genes are a prerequisite for cell death to be executed. Another gene, ced-9, protects against cell death by interacting with ced-3 and ced-4. He has shown how these genes interact with each other in the cell death process and that corresponding genes (a ced-3-like gene) exists in humans. Role of Apoptosis in development of Caenorhabditis elegans The C. elegans genome is 9.7 x 107 bp and is now fully sequenced (24 x E. coli, 1/38 of human). The 19099 genes include 790 seven-pass transmembrane receptors, 480 zinc finger proteins, and 410 protein kinases; see News and Views, Nature 396: 620621 (1998). The life cycle of C. elegans from egg to sexual maturity (and new eggs) is about 3 days. The adult hermaphrodite consists of exactly 959 somatic cells of precisely determined lineage and function. Individual cells are named and their relationships to their neighbours are known. Overall the 959 cells of adult C.elegans arise from 1090 original cells; exactly 131 cells undergo programmed cell death in the wild type worm. Of the 1090, 302 are neurons, and many of the programmed deaths also lie in the neuronal lineage. Identification of Cell Death genes ced-3, ced-4 and ced-9 (C.elegans cell death genes) Robert Horvitz egl-1 (C.elegans egg laying defective). The egl-1 gene was the first gene discovered in the programmed cell death system, as a gain of function mutation causing unscheduled death of two neurons innervating the vulva, and hence the egg laying defect. Ced-4 was then found as the extragenic suppressor of egl-1. Ced-3 suppressed the persistence of cell corpses in phagocytosis defective animals. Ced-1,-2,-5,-6,-7,-10 all turn out to be phagocytosis or clean-up genes rather than acting in the causative pathway of cell death. Ced-3 has a mammalian counterpart, originally known as ICE (Interleukin 1ß Converting Enzyme), now termed Caspase 1 (Cys catalytic Asp targeting protease). Thirteen caspases are known in mammalian systems, and have conserved sequence and subunit structure; of these four play key effector roles in apoptosis and four are initiators in the activation process. Ced-4 acts as an adapter for caspase activation; the mammalian counterpart Apoptosis activating factor Apaf-1. Genetic Mechanism of Apoptosis : The apoptotic system in C.elegans. Meier et.al Nat Reviews, Vol 407, 2000. • Molecular nature of the PCD came from genetic studies done on C.elegans. • 131 cells out of 1090 cells undergo PCD: genetic screens of mutants identified Egl-1, CED3, CED4 and CED 9 to be involved. • Egl-1, CED3, CED4 are death promoters since their loss of function results in survival of all 131 doomed cells. • CED 9 is death inhibitor since its loss of function causes embryonic lethality by massive ectopic cell deaths. Molecular Identities of apoptotic genes Kauffmann and Vaux. Oncogene,22,2003 • Fundamental components of apoptotic pathways are conserved across the species. • Ced -9 is similar to human oncogene :Bcl2. • Ced-3 has a mammalian counterpart, originally known as ICE (Interleukin 1ß Converting Enzyme), now termed Caspase 1. • Ced-4 acts as an adapter for caspase activation; the mammalian counterpart Apoptosis activating factor Apaf-1. Caspases : Cysteine Aspartate Specific ProteASEs • Caspases :more visible "hallmarks" of apoptosis. • Highly specific proteases that cleave proteins exclusively after aspartate residues. • Sequence of 3 amino acids before aspartate determines substrate specificity. • Function: Regulate proteolysis during apoptotic cell death. • Synthesized as inactive zymogens. • Upon activation, twice cleavage at AspX site releases large, small subunits. • Active caspases are tetramers: two large and two small subunits : two active sites. Caspase types: Based on sequence of activation: 1. Initiator (Activator) caspases • First to be activated on commitment of cell to die • Cleave & activate effector caspases • prodomains: Long Contain regulatory sequences • Effector (Executioner) caspases • Cleave & activate cellular substrates • Prodomains: Short. No known regulatory sequences 3. Cytokine processors (Inflammatory) • Prodomains: Long; Contain regulatory sequences •Expressed widely in cells in inactive proenzyme form (pro-caspase) •Must be activated for proteolysis •Responsible for the more visible "hallmarks" of apoptosis •More Notable Members •Caspases 3, 6, 7: Important downstream effector caspases •Caspase 8: initiator in death receptor pathway •Caspase 9: Initiator in intrinsic pathway. Activated by conformational change Effector Caspases: Activate downstream caspases and act on Various cellular substrates In the first example, the effector caspases cleave an inhibitor or an effector protein. An example of this would be CAD (Caspase-Activated Deoxyribonuclease), and ICAD (Inhibitor of CAD). When ICAD binds to CAD, CAD is kept inactivated. However, active effector caspases cleave ICAD which then releases CAD. CAD can then cleave the DNA into fragments (forming the characteristic DNA laddering of apoptotic cells). The second example illustrates that the effector caspases can also cleave structural proteins, such as the nuclear lamins. Nuclear lamins maintain the integrity of the nucleus, but when they are cleaved by the effector caspases, the nucleus condenses (another characteristic of apoptotic cells). Finally, in the third example, effector caspases can cleave off the auto-inhibitory domains of certain proteins. A good example of this would be PAK2. When the effector caspase cleaves off the auto-inhibitory domain of PAK2, PAK2 now becomes constitutively active, playing a role in the membrane blebbing that is characteristic of apoptotic cells. BCL-2 BCL-2 is a human proto-oncogene located on chromosome 18. Its product is an integral membrane protein (called Bcl-2) located in the membranes of the endoplasmic reticulum (ER), nuclear envelope, and in the outer membranes of the mitochondria. The gene was discovered as the translocated locus in a B-cell lymphoma In the cancerous B cells, the portion of chromosome 18 containing the BCL-2 locus has undergone a reciprocal translocation with the portion of chromosome 14 containing the antibody heavy chain locus. This t(14;18) translocation places the BCL-2 gene close to the heavy chain gene enhancer. Bcl-2 Members • Bcl-2 :first protooncogene gene to be discovered from the family: cloned from t(14;18) breakpoint in follicular lymphoma • Presence of Bcl-2 homology domains: BH domains. • BH1 and BH2: in death antagonists, allow heterodimerization with Bax to repress apoptosis • BH3: in death agonists (eg. Bax, Bak) allows heterodimerization with Bcl-XL and Bcl-2 to promote apoptosis • BH4: conserved in apoptosis antagonist members (eg. Bcl-XL) but absent in apoptosis agonists (except Bcl-Xs), this domain allows interaction with death regulatory proteins such as Raf-1, Bad, and perhaps Ced-4. The pro-survival family: • Their hydrophobic carboxy-terminal domain helps target them to the cytoplasmic face of three intracellular membranes. • Bcl2 is an integral membrane protein, even in healthy cells, whereas Bcl-w and Bcl-xL only become tightly associated with the membrane after a cytotoxic signal. • A hydrophobic groove, formed by residues from BH1, BH2 and BH3, can bind the BH3 α-helix of an interacting BH3-only relative. The BH3 only family: • BH3-only proteins seem to be sentinels that are charged with triggering apoptosis in response to developmental cues or intracellular damage. • They are thought to act by binding to and neutralizing their pro-survival relatives. • They act upstream of Bax sub-family of proteins. The Bax family: • Bax is a cytosolic monomer in healthy cells, but it changes conformation during apoptosis, integrates into the outer mitochondrial membrane and oligomerizes. • Whereas, Bak is an oligomeric integral mitochondrial membrane protein, but it too changes conformation during apoptosis and might form larger aggregates. • Bax and Bak oligomers are believed to cause permeabilization of the outer mitochondrial membrane, allowing efflux of apoptogenic proteins, leading to caspase activation. Pro-survival proteins can also act by inhibiting Bax/Bak oligomerization • Heterodimerization of Bcl-2 and Bax inhibits Bax function. • Bcl-XS binding to Bcl-2 prevents Bcl2 from binding to and neutralizing Bax. • Also, heterodimerization with Bax and other pro-apoptotic members of the Bcl-2 family results in the the release of Apaf-1 from Bcl-xL and further activation of caspases. Interactions of Bcl-2 members • Bcl2 family proteins regulate apoptosis via their effect on mitochondria. • Antideath : prodeath molecules e.g Bcl-2/Bax Prodeath: prodeath molecules e.g Bid/Bax Multimerization of same molecules. e.g Bcl2/Bcl2, Bax/Bax. • Either localised in the mitochondria or are induced by death signals. • Translocation to mitochondria is facilitated by post translational modification: Conformation change Caspase cleavage Phosphorylation and dephosphorylaion. Bcl-2 family members can regulate apoptosis related mitochondrial changes such as the ‘permeability transition pore’ (PT) • The PT pore is a poly protein channel, comprising of VDAC and PBR on the outer membrane and ANT and cychlophilin D on the inner membrane. • Cytochrome c release may occur through outer membrane rupture resulting from mitochondrial swelling caused by PT pore perturbation. • Bcl-2, Bcl-xL and Bax have been shown to form ion channels in synthetic lipid membranes. • Thus, Bcl-2 members having the BH1 and BH2 domains may function by forming pores in organelles such as mitochondria or rather, stabilize or perturb the pre-existing channel, PT. Models for release of cyt c from mitochondria • Involves closure of voltage dependent ion channel (VDAC) and impairment of ATP-ADP exchange. • Opening of permeability transition pore (PTP). • Channel formed in the outer mitochondrial membrane By Bax only By Bax and VDAC in combination By lipid or lipid protein complex The Role of Mitochondria in Apoptosis The mitochondrion has been identified as playing a central role in apoptosis 1. Bcl-2 and Bcl-XL localize to the mitochondrial membrane.. 2. Bcl-2 can recruit kinases such as Raf-1 which are involved in mediating Bcl-2's death antagonizing action. 3. Bcl-2/-XL can recruit ced-4 and its mammalian homolg, Apaf-1 to the mitochondrial membrane. This may prevent ced-4/Apaf-1 from activating caspases, thereby inhibiting apoptosis. 4. Mitochondial proteins, when leaked into the cytosol, are capable of inducing apoptosis. During apoptosis, cytochrome c and SMAC are released from the mitochonria and with other factors, such as Apaf-1 (apoptosis protease activating factor-1) and Apaf-3, lead to caspase activation and apoptosis. Increased levels of Bcl-2 can prevent the release of these molecules, whereas, caspase inhibitors cannot. This indicates the release of cytochrome c and SMAC is downstream of Bcl-2 function but upstream of the caspases. 5. Apoptosis is associated with a change in the mitochondrial membrane potential, a phenomenon known as permeability transition (PT). The PT can be blocked by excess Bcl-2 but not by inhibitors of caspases, indicating the PT is downstream of Bcl-2 but upstream of caspase activation. 6. Bcl-2, Bcl-XL, and Bax are capable of forming selective ion pores in membranes. Thus, they may form channels in the mitochondrial membrane that could regulate the PT and the release of molecules such as cytochrome c and AIF. BAD is phosphorylated by Akt, binds to 14-3-3 and is degraded The BAX gene, the promoter of apoptosis, is mutated in genetically unstable cancers of the colorectum, stomach, and endometrium (1998) Inactivating mutation of the pro-apoptotic gene BID in gastric cancer (2004) (6%) Inactivating mutations of proapoptotic Bad gene in human colon cancers. (4.3%) 2004 Caspase-8 gene is frequently inactivated by the frameshift somatic mutation 1225_1226delTG in hepatocellular carcinomas (2005) 10% Somatic mutations of CASP3 gene in human cancers 2004 occasiona Deletion and aberrant CpG island methylation of Caspase 8 gene in medulloblastoma.2004 The Death Receptor Family • Cell surface cytokine receptors belonging to TNF/NGF receptor superfamily. • Receptors are Type I transmembrane proteins: intracellular C terminal tail, membrane spanning region, an extracellular ligand binding domain. • Significant homology in 60-80 aa cytopl. sequence – Death domain (DD). • Death receptors are activated by their natural ligands: group of cytokines belonging to TNF family. Death Receptor Signaling • Ligand binding to the death receptors leads to oligomerization of these receptors. • Recruitment of adaptor molecules through their Death Domains (DD) •This protein –protein interaction is restricted e.g Fas , DR4, DR5 recruits FADD whereas TNFR1 recruits TRADD. • Adaptor molecules recruit initiator caspases through interaction of their Death effector Domain (DED) and CARD domain in caspases. •The resulting complex is called Death Inducing Signaling (DISC) complex. Role of Fas/FasL • Deletion of activated T-cells at the termination of an immune response • Cytotoxic T-cell mediated killing of cells (virus-infected, cancerous) • Destruction of inflammatory or immune cells in immuneprivileged sites (i.e, eyes, reproductive organs) Two pathways in TNFR signaling Pathways to Apoptosis Inhibitor of Apoptosis Family 1993: First member identified in baculoviruses one or more BIR domains critical for activity 1995: first mammalian IAP identified NAIP: positional cloning : spinal muscular atrophy 1997: XIAP, c-IAP1 and 2 shown to inhibit caspase activity 1999: NMR structure of XIAP, caspase binding active site 1999: yeast has IAPs but no caspases: other function Survivin: cell cycle regulation The IAP proteins have been divided into three classes (classes 1, 2, and 3) based on the presence or absence of a RING finger and the homology of their BIR domains. Inhibitors of Apoptosis (IAPs) • IAPs function as intrinsic regulators of caspase cascade to apoptosis. • Inhibit both initiator and effector caspases. • First member identified in baculoviruses: one or more BIR domains critical for activity •IAPs are characterized by 70-80 aa Baculoviral IAP Repeat (BIR) domains. • IAPs with multiple BIR domains use third BIR domain to inhibit caspase 9 and second BIR domain to inhibit caspase 3 ,7. • BIR 1 domain has no caspase inhibiting activity and is least conserved. • RING domain function as adaptors : provide specificity for proteosomal degradation. • NOD domain facilitates self association whereas coiled coil domain mediates interaction with Beta Tubulin. IAP inhibitors : Smac/Diablo, Omi and XAF1. IAPs inhibit active caspases: XIAP inhibits caspases 3, 7 and caspase 9 through separate domains. Its BIR2 domain (amino acids 163–240) with its NH2-terminal extension (amino acids 124–162) inhibits caspases 3 and 7, whereas its BIR3 domain (amino acids 241–356) inhibits caspase 9. These studies provided the basis for developing IAP inhibitors that target the caspasebinding pockets of the molecule. IAP regulators • Regulatory IAP-binding proteins were first identified in Drosophila. The proteins Reaper, Hid, Grim, and Sickle were shown to bind and inhibit the Drosophila IAP, DIAP1. • Later, human IAP inhibitors identified called SMAC/DIABLO and Omi/HTRA2. These IAP inhibitors share a homologous sequence in their NH2 terminus that is responsible for binding and inhibiting IAPs. SMAC and HTRA2 • Human SMAC and HTRA2 are mitochondrial proteins that are released along with cytochrome c during the disruption of the mitochondria. In their active state, SMAC and HTRA2 bind IAPs, thereby preventing their association with caspases. • The IAP-inhibitory functions of the SMAC family of proteins are encoded in their NH2 terminus. Mutation of the NH2-terminal alanine to glycine abolishes the ability of the SMAC peptide to bind IAPs and exert its proapoptotic function. Similar results have been observed with HTRA2. • Structural studies have demonstrated that SMAC binds XIAP at two distinct sites. The NH2 terminus of active SMAC (residues 56–59) binds the BIR3 pocket of XIAP and competitively inhibits the BIR3 domain from binding caspase 9. • SMAC also bind the BIR2 domain of XIAP, but with lower affinity than that for BIR3. The mechanism by which SMAC disrupts the association of BIR2 from caspase 3 is unclear. • HTRA2 binds to the BIR3 domain of XIAP, but with weaker affinity than SMAC. In addition to inhibiting IAPs through binding the BIR3 pocket, HTRA2 can also cleave and inactivate multiple IAPs including XIAP,cIAP1, and cIAP2, but not survivin. Omi has serine protease activity Caspase-independent Apoptosis Death associated with activation of lysosomal and proteosomal Proteases and granzyme B and matrix metalloproteases, calpains. Many programmed or physiological deaths do not appear to depend on caspase activation. Caspase 3 or 9 k. o. embryos die only after embryonic day 10 Techniques to detect Apoptosis Determination of Cell Viability Test : 1) Vital Dye exclusion assay: Trypan blue, propidium iodide : do not stain the viable cells Fluorescein diacetate, Calcein-AM : stain the viable cells. 2) Measurement of cytosolic Leakage: based on the fact that viable cells have intact cellular components. Lactate Dehydrogenase Pyruvate + NADH Lactate + NAD+ NADH NADH exhibits fluorescence at an excitation wavelength of 360 nm with emission at 450nm. Velocity of decrease of Ex 360nm/Em450 nm indicates conversion of NADH to NAD and hence activity of LDH. 3) Clonogenic assay: Ability of cells to divide and form colonies is called clonogenic activit However, integrity of pl memb is not compromised till late stage and hence not very usefu Alterations in Plasma Membrane: • In normal cells, phosphoidylcholine and sphingomyelin are on external leaf and phosphotidylethnolamine and phosphatidylserine (PS) are in the inner leaflet . • Redistribution of phospholipids in the plasma memb. is an early apoptotic change. • Annexin V conjugated with flourophore like FITC can bind to exposed to PS in Ca dependent manner. • Can be viewed microscopically or by flow cytometry. Annexin V staining Alterations in Cytosol: caspase activation • Activation of caspases: hallmark of apoptosis, can be measured. •Western blot for PARP, small subunits of caspase 3,7,8 and 9. • Biochemical analysis of caspase activity: Tetrapeptide sequence (recognition site of caspase) conjugated with report group like p-nitroanilide (pNA) 7- amino -4-methylcoumarin (AMC) 7-amino-4 –triflouromethylcoumarin (AFC) Calorimetric/flourimetric measurements. Immunohistochemica • Immunostaining/flourescent substrates in tissues/cells. l/immunoflorescent staining on paraffin sections. Cell permeable flourescent substrates : PhiphiLux ( Oncogene research products) Bcl2 family Proteins: • Western blot with antibodies specific to individual members. •If subcellular localisation is not relevant: cells can be lysed in non-ionic detergent. • If subcellular is needed, cellular components are subfractionated to obtain mitochondrial and cytosolic fractions. • To determine Bax or Bak oligomers, crosslinking agents are added e.g disuccinimidyl suberate (DSS), Bis (Sulphosuccinimidyl) suberate (BS3) to the mitochondrial fraction suspended in isotonic buffer . • The crosslinker is quenched with 1 M Tris-HCl ph7.5. • Membranes are then lysed in radioimmunol precipitation assay (RIPA) buffer and cleared by centrifugation at 12000g: analysis by SDS-PAGE and Western Blot. • Oligomerisation is visualised using Western blot as a high molecular weight species. MitochondrialChanges: Changes: Mitochondrial Mitochondrial release of Cyt C • Cyt c release is the most common parameter of active mitochondrial pathway. • Subcellular fractionation to yield mitochondrial fraction which is suspended in isotonic buffer with energising agents. • Western Blot of both supernatant and pellet with cyt c antibody: cyt c in sup and reduction in the pellet indicate cyt c release. • ELISA: Supernatant is prepared for ELISA for detection of Cyt c. •Immunostaining cells undergoing apoptosis are washed and fixed Incubated with anti cyt c Ab Wash and incubation with sec Ab tagged with flourophore Diffused cytoplasmic staining indicates cyt c release. Mitochondrial Changes: Mitochondrial Transmembrane Potentials: • Mitochondria transmembrane potential is a functional parameter during apoptosis. •Can be determined by lipophilic cations: accumulation is potential dependent. • Commonly used are : Rhodamine 123 (Rh 123), DiOC6, Tetramethylrhodamine methyl ester (TMRM), JC-1. • Probes can be directly added to cultured cells or isolated mitochondria, incubated for 15 min and harvested to be analyzed by flow or fluorescent microscopy. Changes in the Nucleus: Nuclear Condensation and Fragmentation: • Nuclear condensation and fragmentation can be visualised by staining with fluorescent dyes e.g Hoechst (bisbenzimide) and DAPI (4’6’diamidino 2-phenylindole, dilactate). • Intensity of staining in the nucleus is proportional to the extent of apoptosis due to increased permeability of the dyes. DNA content staining by Propidium Iodide:: Control Tamoxifen Treated Tamoxifen G1 Control S G2 G0 • Degradation of nuclear DNA results in decrease in DNA content or hypoploidy. • Cells are suspended in ice cold PBS, fixed with cold ethanol. • Cells are permeabilised with Triton x100, stained with PI and analysed with flow. • However, necrotic and other cellular debris can also get included. DNA Fragmentation: • Cleavage of DNA at nucleosomal sites results in 180-200 bp fragments which appears as DNA ladder. • Quantitative measurement : amount of fragmented DNA is proportional to frequency of apoptosis. Detection of DNA breaks by TUNEL assay: (Terminal deoxynucleotidyl transferase (TDT) mediated dUTP nick end labeling) • Based on the principle that TdT mediates incorporation of biotinylated dUTP into 3’ OH ends of fragmented DNA . • Cells are permeabilised using Triton X 100 or Proteinase K in case of formalin fixed tissue sections. • Cells/Tissue sections are the incubatd with TdT and dUTPbiotin. • After wash, cells /Tissues can be incubated either with FITC conjugated avidin for detection under fluoroscent microscope or avidin-HRP conjugate can be added followed by DAB. Compounds in development for targeting the BCL-2 family in vivo Compound Genasense Class Antisense oligonucleotide HA14-1 analogs Small molecule Compound 6 Antimycin A3 BH3Is AT101: (–) Gossypol Apogossypol Theaflavanin Polyphenol E GX15-070 ABT-737 Small molecule Small molecule Small molecule Small molecule Small molecule Small molecule Small molecule Small molecule Small molecule IFI-983L, IFI-194 Small molecule CPM-1285 analogs Lipidated peptide Terphenyl derivative Peptidomimetic SAHBs Stapled peptide Mechanism /company Antiapoptotic mRNA downregulation (BCL-2) Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Antiapoptotic inhibition Academic institution stage Genta Developmental Clinical Raylight Chemokine Pharmaceuticals University of Michigan University of Washington Harvard University Ascenta Therapeutics The Burnham Institute The Burnham Institute Mayo Clinic Gemin X Abbott Laboratories/Pfizer (Idun) Infinity Pharmaceuticals/Novartis Raylight Chemokine Pharmaceuticals Yale University Dana-Farber Cancer Institute/Harvard University 4-PhenylsulfanylphenylamineDerivatives Small molecule Proapoptotic inhibition (BID) The Burnham Institute 3,6-Dibromocarbazole Piperazine derivatives of 2-propanol Small molecule Proapoptotic inhibition (BAX) Serono Humanin peptides Peptide Proapoptotic inhibition (BAX) The Burnham Institute Ku70 peptides Peptide Proapoptotic inhibition (BAX) The Blood Center of South Eastern Wisconsin Preclinical Preclinical Preclinical Preclinical Clinical Preclinical Preclinical Preclinical Clinical Preclinical Preclinical Preclinical Preclinical Preclinical Preclinical Preclinical Preclinical Preclinical Thanks