Animated PowerPoint
advertisement

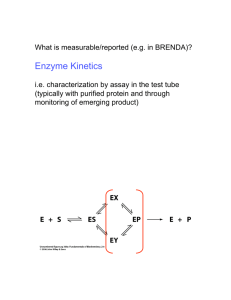
Lecture 15 Chemical Reaction Engineering (CRE) is the field that studies the rates and mechanisms of chemical reactions and the design of the reactors in which they take place. Lecture 15 – Tuesday 3/12/2013 Enzymatic Reactions Michealis-Menten Kinetics Lineweaver-Burk Plot Enzyme Inhibition Competitive Uncompetitive Non-Competitive 2 Review Last Lecture Active Intermediates and PSSH 3 Review Last Lecture Active Intermediates and PSSH 1.In the PSSH, we set the rate of formation of the active intermediates equal to zero. If the active intermediate A* is involved in m different reactions, we set it to: m rA*.net rA*i 0 i 1 2. The azomethane (AZO) decomposition mechanism is k ( AZO) 2 rN 2 1 k ' ( AZO) 4 By applying the PSSH to AZO*, we show the rate law, which exhibits first-order dependence with respect to AZO at high AZO concentrations and second-order dependence with respect to AZO at low AZO concentrations. Enzymes Michaelis-Menten Kinetics Enzymes are protein-like substances with catalytic properties. Enzyme Unease [From Biochemistry, 3/E by Stryer, copywrited 1988 by Lubert Stryer. Used with permission of W.H. Freeman and Company.] 5 Enzymes Enzymes provide a pathway for the substrate to proceed at a faster rate. The substrate, S, reacts to form a product P. S Slow P ES Fast can only catalyze only one reaction. A given enzyme Example, Urea is decomposed by the enzyme urease. 6 Enzymes - Urease A given enzyme can only catalyze only one reaction. Urea is decomposed by the enzyme urease, as shown below. 2O NH2CONH2 UREASE H 2NH3 CO2 UREASE 2O S E H PE The corresponding mechanism is: E S E S k1 E S E S k2 E S W P E k3 7 Enzymes - Michaelis-Menten Kinetics rP k3 E S W rES 0 k1 E S k2 E S k3W E S k1 E S E S k2 k3W Et E E S 8 Et E k1S 1 k2 k3W Enzymes - Michaelis-Menten Kinetics Vmax k3W Et S kcat Et S rP k3 E S W k2 k3W K S M S k1 kcat KM VmaxS rP k3 E S W Km S 9 Enzymes - Michaelis-Menten Kinetics Vmax=kcatEt Turnover Number: kcat Number of substrate molecules (moles) converted to product in a given time (s) on a single enzyme molecule (molecules/molecule/time) For the reaction: kcat H2O2 + E →H2O + O + E 40,000,000 molecules of H2O2 converted to product per second on a single enzyme molecule. 10 Enzymes - Michaelis-Menten Kinetics Michaelis-Menten Equation VmaxS rP rS KM S (Michaelis-Menten plot) 11 Vmax Solving: -rs KM=S1/2 S1/2 CS Vmax VmaxS1/ 2 2 K M S1/ 2 therefore KM is the concentration at which the rate is half the maximum rate. Enzymes - Michaelis-Menten Kinetics Inverting yields: 1 1 KM 1 rS Vmax Vmax S Lineweaver-Burk Plot 1/-rS slope = KM/Vmax 1/Vmax 12 1/S Types of Enzyme Inhibition Competitive E I I E (inactive) Uncompetitive E S I I E S (inactive) Non-competitive E S I I E S (inactive) 13 I E S I E S (inactive) Competitive Inhibition 14 Competitive Inhibition k3 k1 E S E S EP k2 k4 E I E I ( inactive ) k5 1) Mechanisms: E S E S E S P E EI E I rP k 3C ES 15 E S E S E I EI Competitive Inhibition 2) Rate Laws: rES 0 k1CSCE k 2CES k 3CES k1CSC E CSC E C ES k 2 k3 Km k 3CSCE rP Km rIE 0 k 4CICE k 5CIE C I E 16 CICE KI k5 KI k4 Competitive Inhibition C Etot C E C ES C IE k 3C EtotCS rP CI K m K m CS KI VmaxCS rS CI CS K m 1 KI 17 1 1 k m CI 1 1 rS Vmax Vmax K I CS C Etot CE CS C I 1 Km KI Competitive Inhibition From before (no competition): 1 1 K M 1 rS Vmax Vmax CS Increasing C I Competitive No Inhibition 1 rS slope Intercept 18 1 Vmax KM Vmax Competitive 1 1 K M CI 1 rS Vmax Vmax K I 1 CS Intercept does not change, slope increases as inhibitor concentration increases 1 CS Uncompetitive Inhibition 19 Uncompetitive Inhibition Inhibition only has affinity for enzyme-substrate complex E S k1 k2 k3 E S P k4 I E S I E S (inactive) k5 Developing the rate law: rP rS kcat E S rES 0 k1 E S k2 E S kcat E S k4 I E S k5 I E S 20 rIES 0 k4 I E S k5 I E S (1) (2) Uncompetitive Inhibition Adding (1) and (2) k1 E S k2 E S kcat E S 0 k1 E S E S E S k2 kcat KM From (2) I E S k4 I E S I E S I E S k5 KI k5 KI k4 21 kcat E S rp kcat E S KM KI KM Uncompetitive Inhibition Total enzyme Et E E S I E S S I S E 1 KM KI KM kcat Et S rp S I S K M 1 KM KI KM 22 Vmax S rS rP I K M S 1 KI Uncompetitive Inhibition I 1 1 K M S 1 rS Vmax S KI 1 K 1 1 I 1 M rS Vmax S Vmax K I Slope remains the same but intercept changes as inhibitor concentration is increased 23 Lineweaver-Burk Plot for uncompetitive inhibition Non-competitive Inhibition 24 Non-competitive Inhibition E+S +I Increasing I 1 rS -I (inactive)I.E + S Both slope and intercept changes 25 -I P+E +I I.E.S (inactive) rS No Inhibition 1 CS E·S VmaxCS k M CS 1 CI kI 1 1 CI 1 rS Vmax k I kM 1 Vmax CS C I 1 k I Summary: Types of Enzyme Inhibition Lineweaver–Burk plots for three types of enzyme inhibition. 26 End of Lecture 15 27