The StBEL family of transcription factors in potato
advertisement

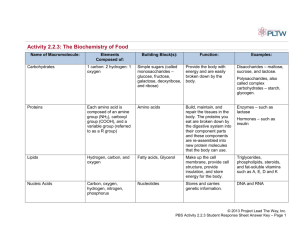
The StBEL family of transcription factors in potato KNOX and BEL1-like proteins are two large families of transcription factors in the three amino- acid loop extension (TALE) superclass (Burglin, et al., 1997). POTH1 (potato homeobox 1) is a member of the knotted1-like family of homeobox genes. During tuber formation, POTH1 plays a positive role by repressing the level of GA in stolon tips (Rosin, et al., 2003). Using the yeast two-hybrid system with POTH1 as the bait, seven distinct BEL1-like proteins (Fig. 1) were isolated from cDNA libraries of potato (Chen, et al., 2003). StBEL genes (13 genes, two are inactive). Based on the whole genome sequence of Solanum phureja in Potato Genome Sequencing Consortium Data, 11 BAC clones were identified with especially low E-values (a good match), using 269 nucleotides from the conserved BELL domain and homeodomain (Fig. 1) of StBEL5 as the query. By HMMBEL1-Like Proteins of Potato based gene structure prediction on SoftBerry website, five new StBEL genes were identified. These are designated StBEL6, -15, -31, -32, -33 and -34. (Fig. 2, StBEL6 is not shown here). Fig. 1 Common structure of BEL1-like proteins Figure 3. A, Schematic of the amino acid sequence of the BEL1-like proteins of potato. Boxed regions represent conserved sequences identified by aligning all seven BELs. Helices I to III of the homeodomain are designated. The Pro-Tyr-Pro (PYP) loop extension is located between helices I and II. For clarity in labeling, the sequence is not drawn to scale. B, Predicted helices of the putative protein-binding region (BELL domain) of the BEL1 protein StBEL5. The bold letters represent amino acids conserved in other plant BEL1 proteins based on a BLAST analysis of StBEL5. The underlined portion of the sequence represents a predicted "-helix. A consensus for the prediction of the sequence structure was derived by using three software programs for amino acid sequence analysis: sspal, ssp, and nnssp (http://www.softberry.com/berry.phtml?topic"protein). Four deletion constructs from Figure 5B are designated with arrows. Construct pAD5-1 contains amino acids 230 through 653 of pAD-05 (interaction with POTH1), and pAD5-2 contains amino acids 257 through 653 of pAD-05 (no interaction). Fig. 2 Phylogenetic tree of the acids BEL1-like proteins Solanum POTH1 is aconsists member of the acids Construct pAD5-11 consists of all amino 1 through 286 ofinpAD-05 (notuberosum. interaction), and pAD5-9 of amino I KNOTTED1-like homeobox 1Class through 315 (interaction with POTH1).genes. POTH1 gives the tree a root. No ESTs were identified for BEL15 and -31. 1. StBEL15 (on St.AC.001.Scaffold000095): Determining the Protein-Binding Regions in the pGAD vector, no interaction was observed with POTH1 and the BEL1-Like Proteins in pBridge. StBEL15 is phylogenetically close to StBEL14, StBEL5 with 49.5% similarity. Fusion constructs of StBEL5 that dissected the 120Interaction with is StBEL5 wasfrom observed all de-StBELs StBEL15 different otherwith identified in the conserved domains. amino acid domain (pAD5-2, -3,First, -4, -9, and -11) were letions outside the KNOX domain, with pBHD2 tested because thisatisthe one of the regions that is conStBEL15 does not have the “SKY” box (Fig.1), with an arginine position for lysine. (missing the amino terminus and the first 48 amino served in BEL TFs from other plant species (Bellaoui StBEL15 lost thewith thirdpBHD6 helix in the homeodomain. The third helix of the acids of Second, the KNOX domain,has Fig. 5A), et al., 2001; Fig. 3B). Interaction with POTH1 was contacts DNA, andofis highly conserved among all the homeobox (missinghomeodomain the carboxy terminus andtarget 52 amino acids observed with all constructs that had deletions excluthe carboxy end of the KNOX domain), and with sively outside of the conserved 120-amino acid box pBHD-9 (first amino-terminal 114 amino acids but no (Fig. 5B). The only exception to this was with KNOX domain sequence). No interaction was obpAD5-9, which demonstrated an interaction and inserved with pBHD3 (missing all of the KNOX docluded a 43-amino acid deletion from the carboxy main and the first 114 amino acids). Control experiend of the 120-amino acid domain. Even with as little ments identified the first 114 amino acids of the N as a 27-amino acid deletion from the amino end of the terminus (pBHD9) as a transcriptional activator. This 120-amino acid domain, interaction did not occur construct transformed alone into AH109 exhibited proteins. StBEL15 does not contain the VSLTLGL box either, normally present in the Ctermini of other StBELs. Besides these changes in the sequence of StBEL15, there are no EST matches with it in the SolEST database. These observations suggest that StBEL15 has lost its function during genome evolution. 2. StBEL31 and StBEL32 (on St.AC.001.Scaffold000399): StBEL31 and StBEL32 are spatially close to each other on the same chromosome, with only 2.2 kb between their respective coding sequences. StBEL31 and StBEL32 are phylogenetically close to StBEL30, with 61.6% and 65.2% similarity respectively. StBEL32 has a similar primary structure to other StBELs, conserved in all the 4 important domains. Blasted in EST database, StBEL32 gets identical matches, indicating that StBEL32 is expressed in potato. Although the protein sequence of StBEL31 is 60% similar to that of StBEL32, StBEL31 is a truncated form of a StBEL gene, missing the VSLTLGL sequence just downstream of Proteins of Potato the homeobox (Fig. 3). Since there is no match for StBEL31 in the EST database,BEL1-Like it is possible that StBEL31 is either nonfunctional or that its mRNA is present at very low levels. Fig. 3 Protein structure of StBEL31 In the genome sequence, StBEL31 is contained in an ORF of a 12.6 kb region, including 23 exons. In the 23 exons, the first five of them encode StBEL31, while the following 14 encode a putative GTP-binding protein, and the last four of them encode a putative peroxidase (Fig. 4). These last two parts have perfect matches respectively in the EST database, meaning that there are two separate proteins encoded by this ORF expressed in potato. The structure of this ORF is like an operon in prokaryotes, which is a cluster of genes operated by a single promoter. However, the three genes are not expressed in the same way, since StBEL31 is not expressed. It’s still unknown how these three genes are controlled separately. Figure 3. A, Schematic of the amino acid sequence of the BEL1-like proteins of potato. Boxed regions represent conserved sequences identified by aligning all seven BELs. Helices I to III of the homeodomain are designated. The Pro-Tyr-Pro (PYP) loop extension is located between helices I and II. For clarity in labeling, the sequence is not drawn to scale. B, Predicted helices of the putative protein-binding region (BELL domain) of the BEL1 protein StBEL5. The bold letters represent amino acids conserved in other plant BEL1 proteins based on a BLAST analysis of StBEL5. The underlined portion of the sequence represents a predicted "-helix. A consensus for the prediction of the sequence structure was derived by using three software programs for amino acid sequence analysis: sspal, ssp, and nnssp (http://www.softberry.com/berry.phtml?topic"protein). Four deletion constructs from Figure 5B are designated with arrows. Construct pAD5-1 contains amino acids 230 through 653 of pAD-05 (interaction with POTH1), and pAD5-2 contains amino acids 257 through 653 of pAD-05 (no interaction). Construct pAD5-11 consists of amino acids 1 through 286 of pAD-05 (no interaction), and pAD5-9 consists of amino acids Fig.4 Structure of the ORF which contains StBEL31 1 through 315 (interaction with POTH1). StBEL33 (on St.AC.001.Scaffold000522) and StBEL34 (on St.AC.001.Scaffold000116): - StBEL33 the andProtein-Binding StBEL34 haveRegions much in lower similarity tothe StBEL5 StBEL15, 31 andwas 32. observed with Determining pGADthan vector, no interaction They are a BEL1-Like separate Proteins branch in the phylogenetic treeStBEL5 of all the StBELs. POTH1 andin the in pBridge. - StBEL33 and StBEL34 are very conserved in the homeobox 97.1% that anddissected the 120Fusion domain, constructswith of StBEL5 Interaction with StBEL5 was observed with all deamino acidhave domain (pAD5-2, -3, -4, -9, and -11) were 95.7% similarity to StBEL5. While, both StBEL33 and StBEL34 lost the SKY box. letions outside the KNOX domain, with pBHD2 (missing the amino terminus and the first 48 amino acids of the KNOX domain, Fig. 5A), with pBHD6 (missing the carboxy terminus and 52 amino acids of the carboxy end of the KNOX domain), and with pBHD-9 (first amino-terminal 114 amino acids but no KNOX domain sequence). No interaction was observed with pBHD3 (missing all of the KNOX do- tested because this is one of the regions that is conserved in BEL TFs from other plant species (Bellaoui et al., 2001; Fig. 3B). Interaction with POTH1 was observed with all constructs that had deletions exclusively outside of the conserved 120-amino acid box (Fig. 5B). The only exception to this was with pAD5-9, which demonstrated an interaction and in- - StBEL34 has extra amino acids at the end of each helix of the BELL domain compared to StBEL5. Especially at the end of the first helix, there are 23 additional amino acids. The similarity between BELL domains in BEL5, -33 and -34: 5 vs. 33 = 72.7%; 5 vs. 34 = 60%; 33 vs. 34 = 59.6%. EST matches have been found for both StBEL33 and StBEL34, indicating that both StBEL33 and StBEL34 can be expressed. 100 AtBEL10 100 AtBLH5 OFP partners AtBLH3 80 StBEL32 StBEL30 72 AtBLH6 98 AtBLH7 99 AtBLH11 63 StBEL31 99 AtBLH8 PNF StBEL34 Shoot Development 100 21 AtBLH9 PNY BLR 40 StBEL33 100 100 StBEL11 33 StBEL5 Tuberization StBEL29 60 AtBLH1 77 StBEL6 Shoot Development AtATH1 100 100 StBEL14 Ovule morphogenesis AtBEL1 StBEL15 63 100 StBEL22 99 StBEL13 Leaf Morphogenesis AtBLH2 SAW1 76 AtBLH4 SAW2 100 AtSTM POTH1 100 99 0.5 Figure 5. Revised phylogenetic analysis of StBEL family in potato reflecting all thirteen genes that have been identified. The amino acid sequence of the thirteen potato BEL-like proteins was analyzed and compared with the thirteen BEL proteins in Arabidopsis. StBELs are represented in bold letters. Putative functions are listed for each group. Because of their presence in the phloem sieve tube system (Fig. 6), their unique UTR sequences and their diverse RNA accumulation patterns, the family of BEL1-like RNAs from potato represents a valuable model for studying the longdistance transport of full-length mRNAs and their role in development. This research will focus on two projects: 1) auto-regulation of the StBEL5 gene and 2) long-distance transport of RNAs of StBEL13, -14 and -30. Fig. 6. RT-PCR products from RNA extracted from either stem sap or stolon tips for several BEL1- like RNAs (designated 5, 13, 14, 22, 29, 30 and 11). Total RNA from Phloem-enriched exudate was extracted and used as the RT-PCR template for detecting tissue-specific transcripts. G2 is RNA for a phloem-specific transcription factor (Zhao et al. 2005) and NT2 is a positive control for root xylem cells (Nazoa et al. 2003). (Campbell et al., 2008). In Arabidopsis thaliana, there are thirteen BEL1-like family members, all of which can form heterodimers with Class 1 KNOX proteins (Hackbusch, et al., 2005; Kumar, et al., 2007). These BEL1-like proteins have significant roles in meristem activities and flower development, and their functions are often overlapping and redundant. BEL1-like proteins in Solanum tuberosum have high homologous relationship with these AtBELs (Fig. 5). StBEL13 matches to SAW1 and SAW2 best. StBEL14 is close to AtBEL1. StBEL5 is close to AtBLH1. StBEL30 is close to AtBLH6 and AtBLH7. ATH1 (ARABIDOPSIS THALIANA HOMEOBOX 1), PNY (PENNYWISE), and PNF (POUNDFOOLISH) are three BEL1-like proteins in Arabidopsis. They are critical for the initiation, maintenance and development of the shoot apical meristem. Triple mutant of these three genes have a more severe phenotype than the single mutants. While in flower formation, ATH1 is a negative regulator, opposite to the role of PNY and PNF (Rutjens, et al., 2009). SAW1 (BLH2) and SAW2 (BLH4) are negative regulators of BP (BREVIPEDICELLUS), an important Class I KNOX gene and positive regulator for growth. In saw1 saw2 double mutant, BP can be expressed on the margin of leaf, and the leaf will grow into a serrated and revolute shape. Since single mutant saw1 and saw2 have no obvious phenotype, the function of SAW1 and SAW2 are redundant in Arabidopsis (Kumar, et al., 2007). AtOFPs are a family of proteins that control the development of ovule in Arabidopsis. Yeast two-hybrid screen revealed complex interaction relationships between AtOFPs proteins and TALE family proteins. Especially, BEL1, BLH1, SAW1, BLH3, SAW2, BLH6 and BLH10 can bind with several of the AtOFPs proteins (Hackbusch, et al., 2005). Indeed, misexpression of BLH1 in embryo sac will switch one of the synergid cells into an egg cell (Pagnussat, et al., 2007), and lose-of-function of BEL1 gene will block the development of integument of ovule (Brambilla, et al., 2007). Do the StBEL proteins also interact with OFPs in potato? Figure 7. Northern-blot analysis of the accumulation of mRNA for four BEL1-like cDNAs (StBEL5, -13, -14, and -30) in potato organs. Ten micrograms of total RNA from flowers, shoot tips (SAM), leaves, stems, roots, unswollen stolons (U stolon), swollen stolons (S stolon), and tubers was loaded per lane. Swollen stolons represent an early stage of tuber formation. A probe for the 18S ribosomal RNA was used to verify equal loading of RNA samples (bottom). A gene-specific probe for each BEL cDNA was used. The length of both 5ʹ′ and 3ʹ′ untranslated sequences are indicated for each BEL RNA. The RNA accumulation patterns of StBEL5, -13, -14 and -30 in different organs has been characterized by Northern blot (Fig. 7). Consistent with the function of AtBEL1-like proteins in ovule development, mRNAs of these four genes accumulate to high levels in flowers, indicating a potential function of StBELs in flower. StBEL5, 13, and 30 also have high mRNA concentrations in SAM, along with the significant role of AtBELs in meristem. StBEL30 has higher accumulation in unswollen stolons, but much lower in swollen stolons and tuber. It is feasible to suggest that StBEL30 may be functional in the tuber induction process. References: Brambilla, V., Battaglia, R., Colombo, M., Masiero, S., Bencivenga, S., Kater, M.M., and Colombo, L. (2007). Genetic and molecular interactions between BELL1 and MADS box factors support ovule development in Arabidopsis. Plant Cell 19, 2544-2556. Burglin, T.R. (1997). Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals. Nucleic Acids Res 25, 4173-4180. Campbell, B., Hallengren, J., and Hannapel, D.J. (2008). Accumulation of BEL1-like transcripts in solanaceous species. Planta 228, 897-906. Chen, H., Rosin, F.M., Prat, S., and Hannapel, D.J. (2003). Interacting transcription factors from the three-amino acid loop extension superclass regulate tuber formation. Plant Physiology 132, 1391-1404. Hackbusch, J., Richter, K., Muller, J., Salamini, F., and Uhrig, J.F. (2005). A central role of Arabidopsis thaliana ovate family proteins in networking and subcellular localization of 3-aa loop extension homeodomain proteins. Proc Natl Acad Sci U S A 102, 4908-4912. Kumar, R., Kushalappa, K., Godt, D., Pidkowich, M.S., Pastorelli, S., Hepworth, S.R., and Haughn, G.W. (2007). The Arabidopsis BEL1-LIKE HOMEODOMAIN proteins SAW1 and SAW2 act redundantly to regulate KNOX expression spatially in leaf margins. Plant Cell 19, 2719-2735. Nazoa, P., Vidmar, J.J., Tranbarger, T.J., Mouline, K., Damiani, I., Tillard, P., Zhuo, D., Glass, A.D., Touraine, B. (2003). Regulation of the nitrate transporter gene AtNRT2.1 in Arabidopsis thaliana: responses to nitrate, amino acids and developmental stage. Plant Mol Biol 52:689–703 Pagnussat, G.C., Yu, H.J., and Sundaresan, V. (2007). Cell-fate switch of synergid to egg cell in Arabidopsis eostre mutant embryo sacs arises from misexpression of the BEL1-like homeodomain gene BLH1. Plant Cell 19, 3578-3592. Rosin, F.M., Hart, J.K., Horner, H.T., Davies, P.J., and Hannapel, D.J. (2003). Overexpression of a knotted-like homeobox gene of potato alters vegetative development by decreasing gibberellin accumulation. Plant Physiology 132, 106-117. Rutjens, B., Bao, D., van Eck-Stouten, E., Brand, M., Smeekens, S., and Proveniers, M. (2009). Shoot apical meristem function in Arabidopsis requires the combined activities of three BEL1-like homeodomain proteins. Plant J 58, 641-654. Zhao, C., Craig, J.C., Petzold, H.E., Dickerman, A.W., Beers, E.P. (2005). The xylem and phloem transcriptomes from secondary tissues of the Arabidopsis root-hypocotyl. Plant Physiol 138:803–818