NATURE –WEB SITE SUPPLEMENTARY INFORMATION for:
advertisement

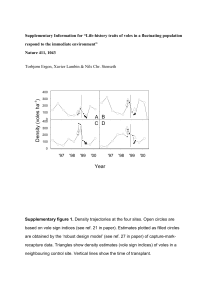
Supplementary Information for “Life-history traits of voles in a fluctuating population respond to the immediate environment” Nature 411, 1043 Torbjørn Ergon, Xavier Lambin & Nils Chr. Stenseth Density (voles ha-1) 400 300 200 100 A B C D 0 400 300 200 100 0 '97 '98 '99 '00 '97 '98 '99 '00 Year Supplementary figure 1. Density trajectories at the four sites. Open circles are based on vole sign indices (see ref. 21 in paper). Estimates plotted as filled circles are obtained by the ‘robust design model’ (see ref. 27 in paper) of capture-markrecapture data. Triangles show density estimates (vole sign indices) of voles in a neighbouring control site. Vertical lines show the time of transplant. Males: 30 25 time *** site *** time × site *** C to D time *** site *** time *** site *** D to A B to A time * site (*) time *** D to B site NS time × site ** B to C Body mass (g) mean ± SE 20 D to C A to D A to B B to D C to A 15 Females: 30 time ** time (*) site ** no change 25 D to C D to A site (*) time ** B to C D to B B to A 20 C to D 15 1/12 1/1 1/2 1/3 C to B A to D 1/12 1/1 1/2 1/3 1/12 1/1 1/2 1/3 1/12 1/1 1/2 1/3 B to D 1/12 1/1 1/2 1/3 Date Supplementary figure 2. Change in body mass of transplanted voles that were recaptured during the first session after transplanting. Lines connect means (±SE) of the same individuals before and after transplant. Changes in individual body mass were analysed with a repeated-measures cross-over analysis (individuals as random subjects and an unstructured covariance matrix for repeated measurements fitted in SAS Proc MIXED28). Significant effects from separate analysis of each pair of transplants for each sex are given in each sub-plot (***: p<0.001, **: 0.001<p<0.01, *: 0.01<p<0.05, (*): 0.05<p<0.1, NS: not selected by the lowest AIC). A ‘time’ effect indicates a change in body mass over time, whereas a ‘site’ effect indicates a change in body mass due to the transplantation between sites. The interaction effect (‘time*site’) implies carry-over effects or effects caused by the fact that the sites were not trapped on the same days. Analysis of transplant success Only 266 of the 761 transplanted voles were ever recaptured after transplant. A potential bias of the differences between transplant groups could arise from differential selection (i.e., transplant success) in the transplant groups. To statistically test whether transplant success was differently related to body mass and reproductive history in the different transplant groups we employed logistic regression models. This analysis show that there is no reason to believe that differential selection among the transplant groups invalidates our interpretation that direct environmental effects are the main source of variation in spring growth and maturation. When considering transplant group (10 groups, see figure 1 in paper), sex, body mass, reproductive history (previously reproduced or not) and all possible interactions, the best model selected by a step-wise AIC-based procedure (the stepAIC function in the S-Plus library MASS (Venables, W. N., and B. D. Ripley. 1997. Modern Applied Statistics with SPlus, Springer)) includes the following effects: Terms added sequentially (first to last) Df Deviance NULL Resid. Df Resid. Dev 758 980.7 Pr(Chi) Group 9 62.64 749 918.1 <0.0001 Sex 1 0.16 748 917.9 0.68 Rep 1 4.84 747 913.1 0.028 Group:Sex 9 32.69 738 880.4 <0.0001 Forcing the Group×Mass effect to stay in the model during the selection procedure gives: Group:Mass Df Deviance 9 5.52 Resid. Df 713 Resid. Dev 848.3 Pr(Chi) 0.79 Forcing the Group×Reproductive history effect to stay in the model during the selection procedure result in: Df Deviance Resid. Df Resid. Dev Pr(Chi) Group:Rep 9 9.98 729 870.4 0.35 Because the tests of the above interaction terms suffer from low statistical power, we also tested the interactions of ‘Site’ and ‘Source’ with ‘Body mass’ and ‘Reproductive history’. The selected model (by the lowest AIC – see above) when considering ‘Site’ and ‘Source’ as predictors includes the following effects: Terms added sequentially (first to last) Df Deviance NULL Resid. Df Resid. Dev 760 985.0 Pr(Chi) Site 3 0.81 757 984.2 0.85 Source 3 46.13 754 938.0 <0.0001 Sex 1 0.09 753 938.0 0.76 Rep 1 4.33 752 933.6 0.037 Site:Source 5 20.46 747 913.1 0.0010 Site:Sex 3 2.92 744 910.2 0.40 Source:Sex 3 20.43 741 889.8 0.0001 Site:Source:Sex 3 9.36 738 880.4 0.025 The ‘site’ and ‘source’ interactions with body mass and reproductive history when these effects are forced to stay in the model during the selection procedure (one by one) results in the following tests: Df Deviance Resid. Df Resid. Dev Pr(Chi) Site:Mass 3 0.494 722 861.5 0.92 Source:Mass 3 1.476 722 860.5 0.69 Site:Rep 3 6.106 737 883.5 0.11 Source:Rep 3 4.338 738 885.4 0.23 In conclusion, there is no significant ‘body mass × transplant group’ or ‘reproductive state × transplant group’ effect on the probability of recapture after transplant. There is a significant effect of reproductive history on transplant success, although there is no significant difference in the relations between the sites. Post-reproductive voles have a lower transplant success than pre-reproductive ones (odds-ratio = 0.7). This probably reflects variation in the “background” mortality rather than responses to the transplant per se because the frequency of post- reproductive voles always decline over the winter (in this and other unpublished datasets from the study system). Although we found no support for differential effects of body mass and reproductive state on the success of transplant, there may still have been differential selection with respect to other unmeasured traits. Nevertheless, two facts point to the importance of individual level responses to changes in the environment even if differential selection has influenced the results: 1. The results presented in figure 2 in the paper clearly show change in individual body mass. 2. The magnitude of the differences in growth and maturation that we observed suggest that selection cannot alone be responsible for the site effects. For example, among the males that originated from site A, even the slowest growing among those moved away from the site grew larger than the fastest growing of the ones that stayed behind in site A. The figure below shows body mass of males that originated from site A. The letter denotes the site the voles lived in (note that most A voles are immigrants, but immigrants were not statistically different from other voles in the same site – see figure 3 in the paper). 50 B B B B B D BB D B 30 Body Mass 40 DD B 20 DD B DD DBB B C C B BB B B A A D DD D DD A A AAAD D A DD A A A A 13 Jan 99 03 Feb 99 24 Feb 99 D DD B D B D D A DD A A A AA CC A AA A AAA AAA A A C D D DDD D AD A AA AA AA A A 17 Mar 99 B DD DD A DD C AA A AAA AA AA AA A 07 Apr 99 B DD A A AAA A AA AAA A AA AAAAA AAA AAA AA AA A A 28 Apr 99 A A A A AA AAA A A 19 May 99 Date We conclude that the interpretation that direct environmental effects are the main source of variation in spring growth and maturation is robust.