VV09
advertisement

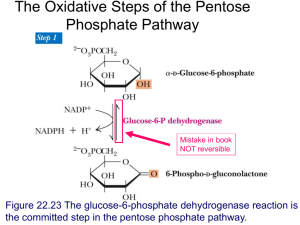
Lecture 9 We started with a bit of a revision of glucose disposal in liver and muscle… but we layered on a couple of extra concepts. 1) GS can be stimulated by a high concentration of G6P – even if the GS is phosphorylated. In other words, a high [G6P] will ‘override’ the inactivating phosphorylation. 2) In the liver, protein phosphatase 1 can be stimulated by a rise in intracellular glucose concentration So this allowed us to see that in MUSCLE, glucose is ‘pulled’ into glycogen by insulin activation of GS (which, in turn, is because insulin stimulates PP1). In contrast, in the liver, glucose is ‘pushed’ into glycogen by the combination of a high [G6P] which stimulates GS, and a glucose-activated PP1. There’s some really good detail on the different ways PP1 is activated in liver and muscle on pages 495-497 of V&V. However, I worry a little about the detail on p486 which gives the impression that the G6P-mediated stimulatory mechanism happens in muscle. I would maintain that the negative feedback of G6P on HK prevents a substantial rise in[G6P] in muscle. I really like the statement in italics regarding metabolic control in section 15-3A. As we’ll see later, the breakdown of glycogen is also regulated by rate limiting enzymes at the level of glycogen… and having rate limiting enzymes responsible for both formation and degradation really allows control DESPITE prevailing substrate concentrations. So then on to describe the molecular detail of glycogenesis and (at least the top bit) of glycolysis. For this we needed to look at some carbohydrate chemistry (much to the dismay of the people in the 2nd row). Important things…(see the first few pages of Chapter 8 of V&V) general structure of monosaccharides ring and open chain forms and the difference between aldoses and ketoses the fact that hexoses are six carbons… and later that three carbons are trioses the fact that changing the orientation around one chiral centre actually changes the properties of the hexose (eg, change at C4 makes the difference b/w glucose and galactose) the formation of an extra chiral centre at C1 when the ring forms (the anomeric centre) the numbering of the carbon atoms the fact that glucose is the only monosaccharide which, when in the ring formation has all the –OH groups facing out ‘equatorially’. Also on this wonderful board we had the first two steps of glycolysis – production of G6P and then isomerisation to fructose 6-phosphate. This then led us onto the next steps – production of F1,6BP by PFK and then the splitting of the hexose phosphates into two triose phosphates. This all reprsents the first stages of glycolysis… where, ironically, all that we’ve done is INVEST ATP… it’s from the G3P and DHAP that we’ll actually make ATP. You’ll find Sections 14-2A (hexokinase), 14-2B phosphoglucose isomerase, 14-2C PFK, 14-2D (aldolase) and 14-2E (triose phosphate isomerase) of interest… although more detail than you ‘need’. I’d be happy if you just knew what was on the board! Note that the board also enabled us to realize that dihydroxyacetone and glyceraldehyde are the simplest ketose and aldose! With the next board, the lecturer in Chemistry that comes after us thought it prudent to correct my poor chemistry by adding Os to the Ps in UTP and UDP-glucose. How we laughed. The jolly japes that we academics enjoy Anyway, this board showed the general strategy of getting glucose into an existing glycogen chain… activation by putting it onto UDP followed by addition to the C4 end of the glycogen molecule (forming an alpha 1 4 glycosidic bond). We saw how the activation step was driven to completion by the formation of pyrophosphate which spontaneously hydrolyses to two phosphates (and essentially renders the reaction irreversible) Next came the silliest board of the semester (or at least it looks like that in retrospect). Still it made the point about glycogenin being at the ‘core’ of every glycogen molecule and how, as the chain elongates, it is branched (through the formation of alpha 16 linkages. Why not check out the preamble to Section 15-2, and then sections 15-2A (on UDP-G pyrophosphorylase, the enzyme that makes UDP-G), section 15-2B on glycogen synthase… but be sure to miss out the box on glycogen storage diseases! Box 15-3 is really great on the branching pattern and the limit to the size of the glycogen molecule. Indeed, pages 486-487 are generally excellent on the whole glycogenin and branching concepts. There are some nice glycogen pictures on pages 219 and 474. And so to the metabolism of fructose… Definitely worth looking at Section 14-5A which is excellent. Note that the metabolism in the liver by-passes the all-important glycolytic control point of PFK.