Cardiovascular system I and 2
advertisement

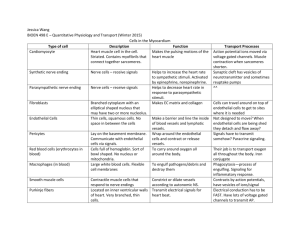
Anatomy and Human Biology 2214 July 21 and 27, 2009 M. Hall CARDIOVASCULAR SYSTEM After studying this lecture, you should be able to: Identify endothelium and understand its important functions Describe the flow of blood through the various chambers of the heart Understand the structure of the heart wall Understand the organization of the impulse conducting system of the heart Identify Purkinje cells and know their function Understand the structure of cardiac muscle Understand the structure of arteries and distinguish between elastic and muscular arteries, small arteries, arterioles and capillaries. What is the functional role of the elastic fibers in elastic arteries? What is the functional role of smooth muscle in muscular arteries? Distinguish between large and small veins and venules Understand the function of the lymphatic system Understand the important role of skeletal and smooth muscle in maintaining venous blood return to the heart. The cardiovascular system consists of the blood and lymphatic vascular systems, and is composed of: The heart--whose function is to pump blood. The arteries--whose function is to carry the blood, with nutrients and oxygen from the heart to the tissues. The capillaries--through whose walls the interchange of nutrients and oxygen between blood and tissues takes place. The veins--whose function is to carry the waste products of metabolism from the tissues to the heart. The lymphatic system--whose function is to return fluid which has leaked into the tissue spaces, to the blood. By distributing hormones, nutrients and oxygen to the cells and tissues of the body, and transporting waste products to the excretory organs, the circulatory system contributes to the integrated functioning of the organism. Endothelium The entire vascular system is lined by endothelium. Endothelial cells are the cells that are in contact with the blood. They are a specialized type of epithelium. Epithelium lines all of the cavities of the body (eg. respiratory system, digestive system, urinary system and the cardiovascular system), as well as the outside surfaces of the body (eg. skin, cornea). Epithelium exists in a number of different forms, which depend on its function. All epithelia sit on a basement membrane, which plays an important role in the function of the epithelium. Underneath this basement membrane is connective tissue, containing collagen and elastic fibers, blood vessels and nerves and various organs such as, in skin, sweat glands, sensory receptors etc. Another important feature of all epithelia is that 1 they have a free surface—one side always faces towards an open space, while the other is attached to the basement membrane. Types of epithelium The endothelial cells that line the entire vascular system are actually just a simple squamous epithelium. They are flattened cells that look like fried eggs. However endothelial cells are not just a physical barrier lining the cardiovascular system. They have many highly specific functional roles, viz: 1. They normally secrete factors which prevent blood clotting (thrombomodulin) 2. They secrete factors that regulate the tone of the vascular smooth muscle (endothelins, nitric oxide, prostacyclin). In this way they are able to regulate the flow of blood through the vessel, by contracting or relaxing the smooth muscle surrounding the vessel. 3. When activated by cytokines, they express cell adhesion molecules which allow white blood cells to stick and migrate into the adjacent connective tissue. 4. In many tissues they actively transport molecules from the blood in the vessel lumen into the tissues that they nourish, by means of pinocytotic vesicles. 5. They function in the formation of new blood vessels (angiogenesis) 6. They contain Weibel-Palade bodies. There are two major constituents of WeibelPalade bodies. One is von Willebrand factor (vWF), a multimeric protein involved in blood coagulation. The second are various selectins which bind to passing immune cells (leukocytes). This allows the fast moving leukocytes in the blood to bind to the endothelial cells lining the blood vessels and slow down in a series of steps called the leukocyte adhesion cascade. The leukocyte adhesion cascade is a sequence of adhesion and activation events that ends with extravasation of the leukocyte, whereby the cell exerts its effects on the inflamed site. At least five steps of the adhesion cascade are capture, rolling, slow rolling, firm adhesion, and transmigration. Subsequently, leukocytes transmigrate across the endothelium (diapedesis) and enter the surrounding tissue where they can migrate to the site of infection. A bewildering number of molecules are involved in this process—the main ones being selectins, chemokines and integrins. 2 Diapedesis 7. In capillaries, endothelial cells take on a number of different configurations depending on the tissue they serve, and the specific functions of that tissue. This will be discussed later. Heart. The heart is a muscular organ that contracts rhythmically, pumping the blood through the circulatory system. It contains four chambers (two atria and two ventricles) through which blood is pumped. Valves guard the exits of the chambers, preventing the backflow of blood. Septae separate the left and right halves of the heart. . The atria receive blood from the body or lungs, while the ventricles pump blood to the body or lungs. For this reason the ventricles are much bigger, and contain more cardiac muscle than the atria. The walls of the heart consist of three layers (tunics), the internal or endocardium which is lined with endothelium; the middle or myocardium; and the external or epicardium which is also lined with a simple squamous epithelium, but here it is called a serosa. Sigh! Layers of the heart: The endocardium is the innermost layer of the heart and is continuous with the t. intima lining the large vessels entering or leaving the heart (the aorta and the venae cavae). It consists of a single layer of endothelial cells resting on a thin basement membrane and a subendothelial layer of varying thickness consisting of loose connective tissue that contains elastic and collagen fibers, as well as some smooth muscle cells. The endothelial cells are in direct contact with blood. Between the endocardium and the myocardium is a layer of connective tissue which contains nerves, veins and Purkinje cells. The myocardium is the middle and thickest of the tunics of the heart and consists of cardiac muscle cells arranged in layers that surround the chambers in a complex spiral. A large number of these layers insert into the fibrous skeleton. The epicardium is the outermost layer of the heart and consists of a layer of mesothelial cells on the outer surface of the heart and some underlying connective tissue. The blood vessels and nerves that supply the heart lie in the epicardium and are surrounded by adipose tissue that cushions the heart in the pericardial cavity. 3 The pericardial cavity is a loose-fitting ‘sac’ consisting of an outer layer of fibrous connective tissue, and an inner serosa, also lined with mesothelium. The cells of the mesothelium secrete the fluid which separates the pericardium from the outer layer of the heart, the epicardium, thus allowing for the smooth movement of the heart during beating. The two mesothelial layers face one another across this fluid layer. Fibrous skeleton. The heart contains a supporting fibrous skeleton (chordae tendinae) which is composed of dense connective tissue with a structure very similar to cartilage. The fibrous skeleton encircles the base of the two arteries leaving the heart and the openings between the chambers. It serves as an attachment for cardiac muscle and for the cuspid valves of the atria and ventricles. It also serves as an attachment site for the semilunar valves of the aorta and the pulmonary artery. Valves. The cardiac valves control the flow of blood from the atria to the ventricles and from the ventricles into the pulmonary artery and the aorta. They consist of a central core of dense fibrous connective tissue, containing both collagen and elastic fibers, lined on both Heart valves and supporting skeleton sides by endothelial layers. The bases of the valves are attached to the fibrous skeleton. 4 Regulation of Heart Rate. Cardiac muscle is capable of contracting in a rhythmic manner without any direct stimulus from the nervous system. In the upper part of the right atrium of the heart is a specialized bundle of modified cardiac muscle cells known as the sinoatrial node (SA node). Acting as the heart's natural pacemaker, the SA node "fires" at regular intervals to cause the heart to beat with a rhythm of about 60 to 70 beats per minute for a healthy, resting heart. The electrical impulse from the SA node triggers a sequence of electrical events in the heart to control the orderly sequence of muscle contractions that pump the blood out of the heart. Because of this function, the SA node is referred to as the pacemaker. The SA node initiates an impulse that spreads along the specialized cardiac muscle fibers/cells of the atria, through the atrioventricular node (A-V node) to the ventricles. The AV node serves as a gate (an "electrical relay station") that slows the electrical current before the signal is permitted to pass down through to the ventricles. This delay ensures that the atria have a chance to fully contract before the ventricles are stimulated. The cells of the AV node give rise to the Bundle of His, which connects with the Purkinje fibers. Purkinje fibers are made up of specialized cardiac muscle cells called Purkinje cells. Purkinje fibers form the Left and Right Bundle Branches, which travel in the subendocardial layers to the tip of the ventricles and then back towards the atria. Contraction is initiated in the atria, forcing blood into the ventricles. Then a wave of contraction in the ventricles begins at the apex of the heart, forcing blood from the heart through the aorta to the body and through pulmonary trunk to the lungs. Although the heart doesn’t require an external stimulus to beat, both the sympathetic and parasympathetic divisions of the autonomic nervous system contribute to the innervation of the heart. Although these nerves do not affect generation of the heartbeat, they do affect rhythm. Stimulation of the parasympathetic system promotes a slowing of the heartbeat, while stimulation of the sympathetic system accelerates the rhythm of the pacemaker. The ventricles of heart have two states: systole (contraction) and diastole (relaxation). Electrical spreading during contraction During diastole blood fills the ventricles and during systole the blood is pushed out of the heart into the arteries. The atria contract anti-phase to the ventricles and chiefly serve to optimally fill the ventricles with blood. These contractions are under the control of the SA and AV nodes. 5 Cardiac muscle is similar to skeletal muscle in many ways, consisting of fibers (cells) that are cross-striated. The fibers of cardiac muscle arise by differentiation and growth of single myoblasts, not by the fusion of many myoblasts as is seen in skeletal muscle. Thus the myofibers of cardiac muscle are much shorter than those of skeletal muscle and have only one or two nuclei, which are located centrally in the cell. Cardiac muscle fibers also branch. The contractile unit is the sarcomere. The cross striations are due to the arrangement of thick myosin and thin actin filaments. The dark band (A-band) consists of myosin filaments with overlapping actin filaments, while the light (I-band) consists largely of actin filaments which anchor into the Z-disks. The changes in the structure of a sarcomere during contraction of cardiac muscle are similar to those in skeletal muscle, as are the biochemical reactions involving the release of Ca2+ from the sarcoplasmic reticulum, attachment of the myosin head to actin, hydrolysis of ATP resulting in a conformational change in the structure of the myosin head, and the power stroke which causes shortening of the sarcomere, and thus muscle contraction. Cardiac Muscle Structure of a sarcomere However, a major difference is seen in the input required to initiate contraction: viz. skeletal muscle contracts only after an external stimulus provided by the motor nerve ending, while cardiac muscle cells possess the ability to contract rhythmically at an intrinsic rate in the absence of an external 6 stimulus. Contraction of the heart results from impulses generated within the heart itself (the SA node). However the autonomic nervous system can modify the rate of heartbeat. Sympathetic stimulation (e.g, from a big fright) increases the heart rate, while parasympathetic stimulation slows it. Thus our hearts beat without us having to remind them to do so. They do this 60-70 times/minute, 24/7 for 70 or more years--they are certainly the hardest working muscles in the body. A structural feature peculiar to cardiac muscle is the intercalated disk (ID). These are seen as cross bands by light microscopy. The intercalated disks are end-to-end and side-to-side junctions between individual, adjacent cardiac muscle cells. Since cardiac muscle cells branch and connect to adjacent cells, the intercalated disks are of different lengths. Intercalated disks have transverse portions that run perpendicular to the long axis of the cell, and lateral portions that run parallel to the long axis of the cells. The transverse portions of intercalated disks contain fascia adherentes and desmosomes. These specialized junctions anchor the muscle fibers together, providing the tight connection that is required for a lifetime of stress during beating. On their lateral surfaces the fibers are connected by gap junctions, which provide electrical coupling between the cardiac muscle cells, thus contributing to a network of communicative pathways over which the overall rhythmic activity of the heart is coordinated. Actin filaments of the last sarcomere of each fiber insert into the intercalated disk. Cardiac muscle cells, like skeletal muscle cells, can only increase by hypertrophy, so that after a heart attack, the damaged portion of the heart cannot regenerate. The damaged tissue is invaded by fibroblasts which form scar tissue to repair the damage. Athletes, particularly long-distance runners, develop large hearts to allow the extra pumping of blood required during prolonged exercise. Structure of intercalated disks cardiac muscle (H&E) Smooth Muscle. Since arteries, and to a lesser extent veins contain layers or bundles of smooth muscle, either in the t. media or t. adventitia, it is helpful to be able to identify this tissue and to understand how it functions in vessel contraction. Smooth muscle is found in situations requiring sustained slow or rhythmic contraction, and is not under voluntary control (i.e, it requires an external neural or hormonal stimulus to contract) 7 Smooth muscle Small Artery Smooth muscle cells are spindle shaped and have a single elliptically shaped nucleus. Each smooth muscle cell is surrounded by an external lamina and they are attached to one another by reticular fibers. Capillaries and nerves run in this CT layer. They may occur singly, in small groups or in large sheets or bundles. When cut in longitudinal section, the nuclei appear elongated, as shown above. However, when cut in cross section, the nuclei appear as round dots. Groups of smooth muscle cells are electrically coupled to one another through gap junctions which allows rapid communication between groups of cells. Thus a sheet of smooth muscle cells will contract tonically by spreading the impulse from cell to cell. The contraction of smooth muscle involves the contractile proteins actin and myosin, but the arrangement of these filaments and the chemical processes which control contraction are quite different from that found in cardiac muscle. The contractile proteins crisscross the cell and are inserted into focal densities that are found attached to the cell membrane. When contraction occurs, the contractile filaments pull on these densities changing the shape of the cell from elongated to round, thus shortening the cell and consequently the bundle of smooth muscle. The Distributing Portion of the Cardiovascular System. Arteries and Veins All blood vessels have a number of structural features in common. They are usually composed of the following layers, or tunics: 1. Tunica Intima. The intima consists of a layer of endothelial cells lining the lumen of the vessel, and lying on a basal lamina. Beneath the endothelium is the subendothelial layer, consisting of loose connective tissue, and sometimes smooth muscle cells. In arteries, the intima is separated from the media by an internal elastic lamina, which contains fenestrae (windows) through which substances can diffuse to the underlying tissue. 2. Tunica Media. The media consists mainly of concentric layers of helically arranged smooth muscle cells, with variable amounts of elastic fibers and elastic lamellae, reticular fibers and ground substance. Smooth muscle cells synthesize this extracellular material. In larger muscular arteries, an external elastic lamina separates the media from the adventitia. 8 3. Tunica Adventitia. The adventitia consists of longitudinally arranged collagen and elastic fibers. The adventitial layer gradually merges with the surrounding connective tissue. In large veins such as the vena cava, bundles of smooth muscle are found in the adventitia. Vasa Vasorum. In large vessels, vasa vasorum (vessels of the vessels) provide nutrients and oxygen to the adventitia and the media, since these layers are too thick to be nourished solely by diffusion from the lumen. Innervation. Most blood vessels that contain smooth muscle in their walls are supplied with a profuse network of unmyelinated sympathetic nerve fibers. Discharge of norepinephrine from these nerves results in vasoconstriction. Arteries. Arteries transport blood from the heart to the tissues. Large arteries resist changes in blood pressure in their initial portions (near the heart) and regulate blood flow in their terminal portions. Arteries are classified according to their size into large elastic arteries, muscular arteries of medium or large diameter and arterioles. In general the walls of arteries are thicker than the walls of veins of the same diameter and contain more smooth muscle. 1). Large elastic arteries. These include the aorta and its large branches. These are also called conducting arteries. They contain a large amount of elastic tissue in the media. The intima consists of the endothelium and a relatively thick subendothelial layer of connective tissue. An internal elastic lamina is present, which delimits the intima from the media. The media consist of layers of concentrically arranged elastic laminae, which contain perforations to allow for diffusion of nutrients from the lumen. Between the elastic laminae are smooth muscle cells, reticular fibers and ground substance. The adventitia consists of dense connective tissue containing elastic and collagen fibers, nerves and blood vessels. 2). Muscular arteries. Most of the named arteries in the body are muscular arteries, which are also called distributing arteries. The intima is similar to that of the elastic arteries and may contain smooth muscle cells. The internal elastic lamina is very prominent. The media contains layers of smooth muscle cells, which decrease in number as the size of the artery decreases. Elastic lamellae, reticular fibers and ground substance are present. An external elastic lamina is prominent in larger muscular arteries. The adventitia consists of collagen and elastic fibers and adipose cells. Lymphatics, nerves and a vasa vasorum are also found in the adventitia. 9 Elastic artery Muscular artery 3). Arterioles. These generally contain 1 to 3 layers of concentrically arranged smooth muscle cells in the media. They contain a prominent internal elastic lamina, but no external elastic lamina. The intima consists of the endothelial cells lying on a thin subendothelial layer. The adventitia is thin. The large elastic conducting arteries serve to transport blood away from the heart. The high content of elastic laminae serve to smooth out the large fluctuations in pressure caused by the heart-beat. During ventricular contraction (systole) the elastic laminae of the conducting arteries are stretched and thus reduce the pressure increase due to the outflow of blood. During ventricular relaxation (diastole), ventricular pressure drops to a low level, but the elastic rebound of these conducting arteries helps maintain arterial pressure. The function of the muscular distributing arteries is to furnish blood to the various organs. The muscle layer in these arteries can control the flow of blood by contracting, or not contracting, in response to local chemical or neural input. In atherosclerosis, lesions form due to focal thickening of the intima due to proliferation of smooth muscle and deposition of cholesterol in smooth muscle cells. These changes may extend to the inner part of the media, and the thickening may become so great as to occlude the vessel. The large elastic arteries of the heart are most prone to atherosclerosis. 10 Capillaries. Capillaries are composed of a single layer of endothelial cells rolled up in the form of a tube. The average diameter of capillaries is 7 to 9 m--about the diameter of an erythrocyte. The external surfaces of these cells usually rest on a basal lamina secreted by the endothelial cells. The total length of capillaries in the body is about 96, 000 km. Endothelial cells lining the capillary are held together by zonula occludens junctions. Thus nutrients must generally be transported through the endothelial cells, not between them. Mesenchymal cells are associated with capillaries and small venules. These cells, called pericytes, have the potential to transform into other cell types, particularly into endothelial cells. Following injury, pericytes proliferate and differentiate to form new blood vessels and connective tissue cells, thus participating in the repair process. They also contain contractile proteins such as actin and myosin, suggesting that they participate in contraction. Capillaries can be grouped into three types: 1). Continuous capillaries, which do not have fenestrae (windows) in their walls. Such capillaries are found in all types of muscle tissue, connective tissue, exocrine glands and nervous tissue. Numerous pinocytotic vesicles are present on both surfaces of capillaries in muscles, where they are responsible for the transport of molecules in both directions across the endothelial cell. By contrast, few or no pinocytotic vesicles are encountered in the continuous capillaries supplying most parts of the nervous system. In nervous tissue, nutrients are transported across the endothelial cells by receptor-mediated processes. This feature accounts in part for the existence of the blood-brain barrier. 2). Fenestrated capillaries, are characterized by the presence of large fenestrae (windows) in the walls of the endothelial cells. These fenestrae are 60 - 80 nm in diameter and are covered by a specialized pore diaphragm that is thinner than a cell membrane. A continuous basal lamina is also present. Fenestrated capillaries are found in tissues where a rapid interchange of substances occurs between blood and tissue, such as the kidney, pancreas, liver, intestine and endocrine glands. Some specialized fenestrated capillaries have no diaphragm covering the openings of the fenestrae. Instead, a very thick basal lamina is present which separates the capillary lumen from specialized epithelial cells (podocytes) which cover the capillary. This type of capillary is characteristic of the glomerulus of the kidney. Classification of capillaries 11 3). Discontinuous (sinusoidal) capillaries have greatly enlarged diameters and follow a tortuous path, which greatly slows the flow of blood. The endothelial wall is discontinuous and the endothelial cells show many fenestrations without diaphragms or basement membrane covering the fenestrae. Sinusoidal capillaries are found mainly in tissues where there is an interchange of blood between the sinusoid and the tissue, such as liver, bone marrow and spleen. Capillary blood flow. Capillaries anastomose freely, forming a rich network that interconnects the arterioles and venules. The arterioles branch into small vessels called metarterioles, which surrounded by a discontinuous layer of smooth muscle. These then branch into capillaries that form a network with a large surface area, to facilitate the exchange of nutrients between the tissues and blood. There is ring of smooth muscle at the point where capillaries originate from the metarteriole. This precapillary sphincter can completely stop the flow of blood to the capillary bed. The entire capillary network does not always function simultaneously. The flow of blood can also be directed through arteriovenous anastomoses which enable the arterioles to empty directly into venules. When vessels of the arteriovenous anastomoses contract, all the blood must pass through the capillary network. When they relax, some blood flows directly to a venule instead of circulating through the capillaries. Flow of blood through a capillary network Capillary circulation is controlled by neural and hormonal stimulation. Because of their thin walls and slow blood flow (0.3 mm/sec, vs. 320 mm/sec in the aorta), capillaries are a favorable place for the exchange of water, solutes and macromolecules between blood and tissues. Functions of capillaries. Capillaries perform at least three important functions: 1). They serve as a selective permeability barrier. Capillaries are the sites at which O2, CO2, substrates, and metabolites are transferred from blood to the tissues, and waste products return to the circulation. Permeability of capillary walls varies with the size and charge of the permeating molecules and with the structure of the endothelial cell. Inflammation can alter the permeability of the junctions between endothelial cells, causing swelling at the site of inflammation. 2). Capillary endothelial cells perform important metabolic functions. They produce substances which have an effect on blood flow, and also break down 12 substances which are produced by other cells. 3). When endothelial cells are damaged or die, the underlying connective tissue becomes a site for the formation of a blood clot (thrombus) which can grow and obstruct blood flow, a potentially life-threatening condition. When endothelial cells are present, they prevent such thrombus formation, and are thus said to exert an antithrombogenic effect. Veins return blood to the heart, aided by the action of both skeletal and smooth muscle contraction and by the presence of specialized valves. They vary in size from less than 1 mm to 4 cm in diameter. In comparison to arteries, they have a larger lumen and relatively thinner wall. They are classified according to their size as large, medium or small veins, or as venules, Large vein Medium vein Large veins have a well developed intima, with a subendothelial layer of connective tissue and some smooth muscle cells. The media is thinner than in similarly sized arteries, with a few layers of smooth muscle cells and connective tissue. A distinguishing feature of large veins is the presence of longitudinal bundles of smooth muscle in the tunica adventitia, as well as the presence of collagen and elastic fibers. With the exception of the main trunks, such as the vena cavae, and other named veins (pulmonary, renal, portal etc.) most veins are medium-sized or small. The intima usually has a thin subendothelial connective tissue layer, while the media contains small bundles of smooth muscles, reticular and elastic fibers. The adventitia consists of collagen bundles. Medium and small veins have a similar histology, and vary mainly in the diameter of their lumen, and the amount of smooth muscle in the tunica media and tunica adventitia. Venules have very thin walls. The intima consists of only endothelial cells sitting on a media which contains only a few smooth muscle cells. The adventitia is thin and consists of collagen and elastic fibers. Large and medium veins have valves in their lumens. These are an extension of the intima. They are composed of elastic connective tissue and are lined on both sides by endothelium. The valves direct the flow of blood to the heart, often against the forces of gravity. When the skeletal muscle surrounding a vein is relaxed both the upper and lower valves are closed to prevent backflow of the blood within the vein. As the skeletal muscle on either side of the vein contracts it squeezes the vein putting pressure on the blood inside. This is aided by the contraction of smooth muscle in the adventitia. The blood is pushed both 13 downwards and upwards. Pressure of the blood on the valve causes the lower valve to close and thus no blood can be pushed downwards. However the increased pressure will cause the upper valve to open and allow blood to go up into the next section of the vein. It is because of this important role of skeletal muscles in aiding blood flow that blood pools in the extremities after long periods of inactivity eg. sitting in a plane or in bed-ridden patients. Lymphatic Vascular System. Fluids are constantly exchanged between the capillaries and the surrounding tissues. About 90% of this fluid is returned to the capillary at its venous end. The function of the lymphatic system is to collect the remaining 10% of this fluid from the tissue spaces and return it to the blood. Unlike the blood, it circulates in only one direction---toward the heart. The lymphatic capillaries originate in the various tissues as thin, blind-ended vessels that consist of a single layer of endothelium. These capillaries are held open by numerous elastic fibers, which also bind them firmly to the surrounding connective tissue. Lymphatic capillaries absorb some of the electrolytes and proteins that continuously leave the blood capillaries, and are not reabsorbed. Since there are no tight junctions between the endothelial cells, tissue fluid can easily enter the lymphatic system. The thin lymphatic vessels gradually converge and end up as two large trunks--the thoracic duct and the right lymphatic duct, which enter the venous system. Interposed in the path of the lymphatic vessels are numerous lymph nodes. During passage through the lymph nodes, foreign antigens that have found their way into the lymph can be processed by the immune system. Activated lymphocytes, which have escaped from the blood to enter the connective tissues, and which are important in the immune defense, are added to the lymph. The larger lymphatic vessels have a structure similar to veins, although with thinner walls which contain some smooth muscle bundles. They have internal valves which assure the unidirectional flow of lymph. Lymphatic circulation is aided by the contraction of the surrounding skeletal and smooth muscles. The movement of fluid out of or into capillaries is determined by the difference between the osmotic pressure of the blood and the blood pressure. Thus fluids leave the capillary at the arterial end and reenter the capillary at the venous end. About 90% of the fluid leaving the capillary is returned at the venous end. The remaining 10% enters the lymphatic capillary system. The following link shows a cool animation of this process: http://highered.mcgraw-hill.com/sites/dl/free/0072464631/291136/Fluidexchange.swf 14 Distribution of blood in the body. At any one time, over half of the total blood in the vascular system is in the veins. The remainder is fairly evenly distributed between the other components of the circulatory system. Heart (diastole) Lungs Large arteries Small arteries and arterioles Capillaries Veins 7% 9% 7% 8% 5% 64% 15