23.Acquired Immunity - Dinman, Jonathan D.
advertisement

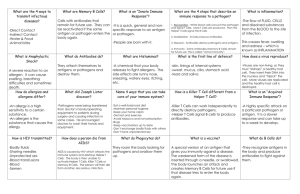
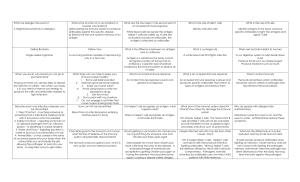
Lecture 23: Virus offence meets host defense: acquired immunity and immunopathology BSCI437 Flint et al. Chapter 15, pp. 531 – 584. Adaptive Immunity is a specific response to a “foreign substance” Specificity: protection is directed against specific foreign molecules. Memory: after initial exposure to foreign antigen, the body reacts more vigorously and quickly at a later meeting with same antigen. Two Branches of the Specific Immune Response 1. Antibody-Mediated Immune Response aka Humoral immunity a specific defense provided by the antibodies targets bacteria, free viruses, fungi, protists 2. Cell-Mediated Immune Response aka Cellular immunity a specific defense mounted by T-cells targets infected cells, cancer cells, transplants Components of the adaptive immune response 1. Antigens – small molecules sensed by… 2. B-cell receptors (a.k.a Antibodies) 3. T-cell receptors Presented in Context of “Self” by cells 1. Macrophages etc…Antigen Presenting Cells 2. Infected (non-self) cells …to cells 3. B-cells – humoral immunity. Mature in bone marrow (?) 4. T-cells – cell mediated immunity. Mature in thymus gland Antigen vs epitope Antigen: antibody generating. A foreign (not “self”) substance that can stimulate an immune response against it. Usually proteins or polysaccharides big, high MW (>10,000 daltons) Epitope – a part of the antigen against that is specifically recognized at the molecular level. An antigen can contain multiple epitopes. Given the repetitive nature of viral capsids and envelopes, each virus tends to display tens (to hundreds depending on T number) epitopes. B- and T- cells have receptors for antigens. B-cell receptor = surface-associated antibody T-cells display a heterodimeric membrane associated T-cell receptor (TCR) Highly specific Interactions between receptors and epitopes are highly specific. One epitope - one receptor B-Cells: Antibodies and Humoral Immunity Antibodies = Immunoglobins. Made by B-cells Immunoglobulins (Ig’s) -- protein molecules produced by B-cells. Basic structure: o 4 chains -- 2 identical heavy chains + 2 identical light chains o Each chain contains N-terminal variable and C-terminal variable regions Antibodies Have 2 Functional Ends: Fab and Fc o Fab: antigen binding region: variable. o Fc: constant. Interacts with Fc receptors on T-cells, macrophages, dendritic cells, mast cells, glial cells, neutrophils, basophils and eosinophils. Can also fix complement. Protective Outcomes of Antibody – Antigen Binding Antibodies bind to specific antigens. Tag them as foreign, for elimination via complement fixation, phagocytosis, cytotoxic clearing, etc. Antibodies also stimulate T-cells and macrophages to produce cytokines. 5 Different Classes (isotypes) of Antibodies (Table 15.13) IgM: Primary antibody. Expressed on surface of B-cell, can be secreted after stimulation. Pentavalent. IgD: A very minor primary antibody. Monovalent. IgG: The major circulatory species. 5 subclasses IgA: The major secertory species. Bivalant. 2 subclasses IgE: associated with mast cells in epithelia. Histamine response, allergies. Natural Antibody Humans and higher primates don’t make galactose (1,3)-galactose (-gal) Almost everything else on the planet does Constant exposure to bacterial -gal High levels of anti-a-gal antibodies An explanation for why we generally don’t get infected by enveloped animal viruses. The Cellular immune response. Involves T-cells T-cell receptors are antigen specific. T-cell receptors recognize complexes of antigen bound to either MHC Class I or Class II molecules. CD4 vs/ CD8 T-cells. Figures 15.19 – 15.21 CD4 and CD8 are co-receptors expressed on the surfaces of T-cells. CD4: associated with ‘helper’ T-cells. o These interact with MHC Class II/antigen complexes on Antigen Presenting Cells (e.g. Macrophages, Dendritic cells, B-cells). o Stimulate and direct the immune response through release of cytokines. CD8: Associated with ‘cytotoxic’ T-cells. Interact with MHC Class I/antigen complexes on somatic cells. o Recognize cells as ‘non-self’, i.e. defective/infected, and directly kill those cells. Virology and the adaptive response: Humoral arm o Fab regions of antibodies recognize specific molecular shapes. o Fc regions o Act as ligands for Fc receptors on phagolytic cells, e.g. macrophages, neutrophils o Can fix complement which can poke holes in viral envelopes. o Can neutralize viral receptors or viral uncoating. o Aggutinate/precipitate viruses Fig. 15.24 shows how these properties of antibodies can work to inhibit infection by Human Rhinovirus. o Virus responses: o Subject receptors to high rates of mutation, i.e. evolve away from the immune response. Examples: HIV-1 Env proteins, Antigenic drift/antigenic shift in haemaglutinnin and neuraminidase proteins in Influenza o Use the antibody as a means of entry into target cell. e.g. Dengue and Yellow fever: Typically, the first infection is mild, and serotype-specific antibodies are produced. Upon subsequent infection with a different serotype, the Fab portion of the antibodies bind virus particle but do not neutralize it. The Fc portion interacts with Fc receptors on macrophages, the infectious target of the virus. Thus the virus gains entry into the macrophage by receptor-mediated endocytosis. Virology and the adaptive response: Cellular arm Expression of viral antigens on the surface of cells in association with MHC Class I marks the cell as infected Targets cell for death by cytotoxic T-cells (CTL). Virus responses: o Downregulate MHC Class I expression – many examples o Infect the T-cells themselves, e.g. HIV-1 o Prevent CTLs from proliferating, e.g. HIV-1 Nef, Tat and Fas; CMV makes at least 6 proteins that promote CTL apoptosis. o Produce virokines to downregulate immune response (Table 15.6) o Produce Viroceptors to divert interferons, interleukins, and other chemokines from their targets (Table 15.7) Immunopathology: too much of a good thing. Pathology is damage to cells and tissues. Viruses can only do this directly if they lyse infected cells. Most viral infections are not lytic however. So how then do viral infections result in pathology? In the end, eliminating virus from the body involves clearing out infected cells. To do this, effector cells are recruited to the site of infection by cytokines, resulting in inflammation. Inflammation provides integration and synergy with the main components of the immune system (Fig. 15.12). Lesions caused by Cytotoxic Lymphocytes Tissue damage requires CTLs. e.g. Hepatitis B. Envelope protein expressed on surface of infected cells. CTLs recognize this in context of MHC class I and induce aptosis by TNF- etc. Other cytokines recruit neutrophils and monocytes which cause more extensive damage. Lesions caused by CD4+ (helper) T-cells CD4+ T-cells produce even more cytokines than do CTLs and recruit and activate many nonspecific effector cells. Such inflammatory reactions are usually called delayed type hypersensitivity responses. Most recruited cells are neutrophils and mononuclear cells. These cells tend to protect against cytopathic viruses. However, with persistent viruses, the result is immunopathological. Immunopathology is the result of release of proteolytic enzymes, reactive free radicals e.g. peroxide, nitric oxide, and cytokines, e.g. TNF-. Immune complex diseases (Fig. 15.25). Virus-antibody complexes can accumulate to high concentrations, especially when infection occurs at sites that are not readily available to immune cells. The resulting complexes are not efficiently cleared, and circulate in the blood. These get stuck in small capillaries, causing lesions that are exacerbated when complement is activated. Common sites of ICD are kidneys (glomerulonepheitis), brain (encephalitis, confusion), lungs and extremities. Autoimmune diseases. Caused by immune response directed against host tissues. Two mechanisms. 1. Viral infection exposes “sequestered antigens”, i.e. proteins that are usually hidden (inside the cell, or in a compartment that is not monitored by the immune system) to the immune system. e.g. Coxsackie virus infection lyses cardiac muscles exposing cardiac myosin to immune system production of anti-cardiac myosin antibodies heart disease. 2. Molecular mimicry: Viral proteins have epitopes that look like those on host cell proteins. Antibodies directed against viral proteins cross react with host proteins. E.g. Lymphatic coriomeningitis virus can induce diabetes mellitis or demyelinating disease in the nervous system in experimental mouse models. Free Radical Induced Immupathology (Fig. 15.26) Superoxide (O2•) and nitric oxide (NO) are produced during the inflammatory response. Interaction of the two produced peroxinitrite (ONOO•) which kills lyses cells. Typically, mononuclear cells are recruited to the site of infection by cytokines, and use (ONOO•) to kill the infected cell. This is a “dumb bomb” strategy however…neighboring cells are killed too. This causes more inflammation, etc.