Plants and their Structure

General Plant Organization
A plant has two organ systems: 1) the shoot system, and 2) the root system. The shoot system is above ground and includes the organs such as leaves, buds, stems, flowers (if the plant has any), and fruits (if the plant has any). The root system includes those parts of the plant below ground, such as the roots, tubers, and rhizomes.
Major organ systems of the plant body
Figure 1
.
Plant cells are formed at meristems, and then develop into cell types which are grouped into tissues. Plants have only three tissue types: 1) Dermal; 2) Ground; and
3) Vascular. Dermal tissue covers the outer surface of herbaceous plants. Dermal tissue is composed of epidermal cells, closely packed cells that secrete a waxy cuticle that aids in the prevention of water loss. The ground tissue comprises the
bulk of the primary plant body. Parenchyma, collenchyma, and sclerenchyma cells are common in the ground tissue. Vascular tissue transports food, water, hormones and minerals within the plant. Vascular tissue includes xylem, phloem, parenchyma, and cambium cells.
Two views of the structure of the root and root meristem. Images from Purves et al.,
Life: The Science of Biology, 4th Edition, by Sinauer Associates
( http://www.sinauer.com/ ) and WH Freeman ( http://www.whfreeman.com/ ), used with permission.
A Plant cell types rise by mitosis from a meristem which is defined as a region of localized mitosis. Meristems may be at the tip of the shoot or root (a type known as the apical meristem) or lateral, occurring in cylinders extending nearly the length of the plant. A cambium is a lateral meristem that produces (usually) secondary growth.
Secondary growth produces both wood and cork (although from separate secondary meristems).
Parenchyma
A generalized plant cell type, parenchyma cells are alive at maturity. They function in storage, photosynthesis, and as the bulk of ground and vascular tissues. Palisade parenchyma cells are elogated cells located in many leaves just below the epidermal tissue. Spongy mesophyll cells occur below the one or two layers of palisade cells.
Ray parenchyma cells occur in wood rays, the structures that transport materials laterally within a woody stem. Parenchyma cells also occur within the xylem and phloem of vascular bundles. The largest parenchyma cells occur in the pith region, often, as in corn ( Zea ) stems, being larger than the vascular bundles. In many prepared slides they stain green.
Schematic diagram of a cross-section of a leaf
Palisade cell
Phloem
Phloem cells conduct food from leaves to rest of the plant. They are alive at maturity and tend to stain green (with the stain fast green). Phloem cells are usually located outside the xylem. The two most common cells in the phloem are the companion cells and sieve cells. Companion cells retain their nucleus and control the adjacent sieve cells. Dissolved food, as sucrose, flows through the sieve cells.
Xylem cells. The above image (left) is from gopher://wiscinfo.wisc.edu:2070/I9/.image/.bot/.130/Stem/
Zea_cross_section/Vascular_Bundle_labelled . The above image (right) is from http://www.biosci.uga.edu/almanac/bio_104/notes/apr_9.html
.
Tracheids, longer, and narrower than most vessels, appear first in the fossil record.
Vessels occur later. Tracheids have obliquely-angled endwalls cut across by bars.
The evolutionary trend in vessels is for shorter cells, with no bars on the endwalls.
Conducting cells of the xylem; tracheids (left) are more primitive, while the various types of vessels (the other three) are more advanced. Image from Purves et al., Life:
The Science of Biology, 4th Edition, by Sinauer Associates ( http://www.sinauer.com/ ) and WH Freeman ( http://www.whfreeman.com/ ), used with permission.
Conductive Vessel Element in Mountain Mahogany Wood (SEM x750). This image is
copyright Dennis Kunkel at http://www.denniskunkel.com/ , used with permission.
.
Phloem cells. The above (left) image is cropped from gopher://wiscinfo.wisc.edu:2070/I9/.image/.bot/.130/Cells_and_Tissues/Cucurbita_Stem/Cross_Section/Phloem/Sieveplate
Cross_Section/Phloem/Sieve-plate . The above (right) image is from http://www.biosci.uga.edu/almanac/bio_104/notes/apr_9.html
.
Phloem cells as seen in longitudinal section. Note the longitudinal view of the sieve plate inside the large sieve tube cell. Right image is a diagram of the longitudinal view of phloem cells. The above image(left) is cropped from gopher://wiscinfo.wisc.edu:2070/I9/.image/.bot/.130/Cells_and_Tissues/
Cucurbita_Stem/Longitudinal_Section/Sieve-plate_l.s
. Right image is from Purves et al.,
Life: The Science of Biology, 4th Edition, by Sinauer Associates ( http://www.sinauer.com/ ) and WH Freeman ( http://www.whfreeman.com/ ), used with permission.
Epidermal Cells
Epidermis
The epidermal tissue functions in prevention of water loss and acts as a barrier to fungi and other invaders. Thus, epidermal cells are closely packed, with little intercellular space. To further cut down on water loss, many plants have a waxy cuticle layer deposited on top of the epidermal cells.
Guard Cells
To facilitate gas exchange between the inner parts of leaves, stems, and fruits, plants have a series of openings known as stomata (singular stoma). Obviously these openings would allow gas exchange, but at a cost of water loss. Guard cells are bean-shaped cells covering the stomata opening. They regulate exchange of water vapor, oxygen and carbon dioxide through the stoma.
Scanning electron micrograph of Equisetum (horsetail or scouring rush) epidermis. Note the oval stomatal apparatuses in the center of the stem. The above image is from http://www.mcs.csuhayward.edu/sem/images/horsel4.gif
.
Epidermal cells, including guard cells, of corn. The above image is from gopher://wiscinfo.wisc.edu:2070/I9/.image/.bot/.130/Leaf/Corn_epidermal_peel .
Pea Leaf Stoma, Vicea sp. (SEM x3,520). This image is copyright Dennis Kunkel at http://www.denniskunkel.com/ , used with permission.
Palisade cells are a type of leaf tissues and can be found within the mesophyll in leaves of dicotyledonous plants. They contain chloroplasts , which convert the energy stored in photons to chemical energy through photosynthesis .
Palisade cells show various adaptations: first, their cylindrical shape, which allows maximum absorption of light by chloroplasts . Second, these cells produce carbohydrates in greater quantities than are needed by each cell; these carbohydrates are fed into a wide variety of metabolic pathways and are vital to the functioning of the plant. Third, the palisade mesophyll contains the largest per-cell number of chloroplasts in the plant; usually positioned towards the upper surface of the leaf in order to harness the greatest amount of energy possible.
Palisade cells are placed all over the top of a leaf, maximising the extent of light absorption.
As a result, the top of a leaf is often darker than the bottom.
Palisade cells are grouped together to give the palisade layer of the leaf- this is the leaf tissue where most of the photosynthesis takes place.
Chloroplast ultrastructure:
1. outer membrane
2. intermembrane space
3. inner membrane (1+2+3: envelope)
4. stroma (aqueous fluid)
5. thylakoid lumen (inside of thylakoid)
6. thylakoid membrane
7. granum (stack of thylakoids)
8. thylakoid (lamella)
9. starch
10. ribosome
11. plastidial DNA
12. plastoglobule (drop of lipids)
Chloroplasts are observable morphologically as flat discs usually 2 to 10 micrometer in diameter and 1 micrometer thick. In land plants they are generally 5 μm in diameter and 2.3
μm thick. The chloroplast is contained by an envelope that consists of an inner and an outer phospholipid membrane. Between these two layers is the intermembrane space. A typical
[parenchyma] cell contains about 10 to 100 chloroplasts.
The material within the chloroplast is called the stroma, corresponding to the cytosol of the original bacterium, and contains one or more molecules of small circular DNA. It also contains ribosomes , although most of its proteins are encoded by genes contained in the host cell nucleus, with the protein products transported to the chloroplast.
Within the stroma are stacks of
thylakoids
, the sub-organelles which are the site of photosynthesis. The thylakoids are arranged in stacks called grana (singular: granum). A thylakoid has a flattened disk shape. Inside it is an empty area called the thylakoid space or lumen. Photosynthesis takes place on the thylakoid membrane; as in mitochondrial oxidative phosphorylation, it involves the coupling of cross-membrane
fluxes
with
biosynthesis
via the dissipation of a proton electrochemical gradient. Help us improve Wikipedia by supporting it financially .
Thylakoid
Thylakoids (green) inside a chloroplast
Thylakoids (green) inside a cyanobacterium ( Synechocystis )
TEM image of the cyanobacterium Prochlorococcus showing the thylakoid membranes
A Thylakoid is a membrane-bound compartment inside chloroplasts . They are the site of the light-dependent reactions of photosynthesis . The word "thylakoid" is derived from the Greek thylakos , meaning "sac". Thylakoids consists of a thylakoid membrane surrounding a thylakoid lumen . Chloroplast thylakoids frequently form stacks of disks referred to as
"grana" (singular: granum ). "Grana" is Latin for "stacks of coins". Grana are connected by intergrana or stroma thylakoids, which join granum stacks together as a single functional compartment. o
[
edit
] Thylakoid structure
Thylakoid structures
Thylakoids are membrane-bound structures embedded into the chloroplast stroma .
Membrane
The thylakoid membrane is the site of the light-dependent reactions of photosynthesis with the photosynthetic pigments embedded directly in the membrane. It is an alternating pattern of dark and light bands mesasuring each 0.001 μm.
[1] The thylakoid lipid bilayer shares characteristic features with prokaryotic membranes and the inner chloroplast membrane. For example, acidic lipids can be found in thylakoid membranes, cyanobacteria and other photosynthetic bacteria and are involved in the functional integrity of the photosystems.
[2]
The thylakoid membranes of higher plants are composed primarily of phospholipids
[3]
and galactolipids that are asymmetrically arranged along and across the membranes.
[4]
The lipids for the thylakoid membranes are synthesized in a complex pathway involving exchange of lipid precursors between the endoplasmic reticulum and inner membrane of the plastid envelope and transported from the inner membrane to the thylakoids via vesicles.
[5]
Lumen
The thylakoid lumen is the compartment bounded by the thylakoid membrane . It plays a vital role for photophosphorylation during photosynthesis . During the light-dependent reaction, protons are pumped across the thylakoid membrane into the lumen making it acidic down to pH 4.
Granum
A granum (plural grana ) is a stack of thylakoid discs. Chloroplasts can have from 10 to 100 grana. Grana are connected by stroma thylakoids, also called intergrana thylakoids or
lamellae . Grana thylakoids and stroma thylakoids can be distinguished by their different protein composition.
Thylakoid formation
Chloroplasts develop from proplastids when seedlings emerge from the ground. Thylakoid formation requires light. In the plant embryo and in the absence of light, proplastids develop into etioplasts that contain semicrystalline membrane structures called prolamellar bodies.
When exposed to light, these prolamellar bodies develop into thylakoids. This does not happen in seedlings grown in the dark, which undergo etiolation . An underexposure to light can cause the thylakoids to fail. This causes the chloroplasts to fail resulting in the death of the plant.
Thylakoid formation requires the action of vesicle-inducing protein in plastids 1 (VIPP1).
Plants cannot survive without this protein, and reduced VIPP1 levels lead to slower growth and paler plants with reduced ability to photosynthesize. VIPP1 appears to be required for basic thylakoid membrane formation, but not for the assembly of protein complexes of the thylakoid membrane.
[6]
It is conserved in all organisms containing thylakoids, including cyanobacteria,
[7]
green algae, such as Chlamydomonas ,
[8]
and higher plants, such as
Arabidopsis .
[9]
Thylakoid isolation and fractionation
Thylakoids can be purified from plant cells using a combination of differential and gradient centrifugation .
[10]
Disruption of isolated thylakoids, for example by mechanical shearing, releases the lumenal fraction. Peripheral and integral membrane fractions can be extracted from the remaining membrane fraction. Treatment with sodium carbonate (Na
2
CO
3
) detaches peripheral membrane proteins , whereas treatment with detergents and organic solvents solubilizes integral membrane proteins .
Thylakoid proteins
Thylakoid disc with embedded and associated proteins shanyn rules almo is so hot!!!!!!
Thylakoids contain many integral and peripheral membrane proteins, as well as lumenal proteins. Recent proteomics studies of thylakoid fractions have provided further details on the protein composition of the thylakoids.
[11]
These data have been summarized in several plastid protein databases that are available online.
[12][13]
According to these studies, the thylakoid proteome consists of at least 335 different proteins.
Out of these, 89 are in the lumen, 116 are integral membrane proteins, 62 are peripheral proteins on the stroma side, and 68 peripheral proteins on the lumenal side. Additional lowabundance lumenal proteins can be predicted through computational methods.
[14][10]
Of the thylakoid proteins with known functions, 42% are involved in photosynthesis. The next largest functional groups include proteins involved in protein targeting , processing and folding with 11%, oxidative stress response (9%) and translation (8%).
[12]
Integral membrane proteins
Thylakoid membranes contain integral membrane proteins which play an important role in light harvesting and the light-dependent reactions of photosynthesis. There are four major protein complexes in the thylakoid membrane:
Photosystems I and II
Cytochrome b6f complex
ATP synthase
Photosystem II is located mostly in the grana thylakoids, whereas photosystem I and ATP synthase are mostly located in the stroma thylakoids and the outer layers of grana. The cytochrome b6f complex is distributed evenly throughout thylakoid membranes. Due to the separate location of the two photosystems in the thylakoid membrane system, mobile electron carriers are required to shuttle electrons between them. These carriers are plastoquinone and plastocyanin. Plastoquinone shuttles electrons from photosystem II to the cytochrome b6f complex, whereas plastocyanin carries electrons from the cytochrome b6f complex to photosystem I.
Together, these proteins make use of light energy to drive electron transport chains that generate a chemiosmotic potential across the thylakoid membrane and NADPH , a product of the terminal redox reaction. The ATP synthase uses the chemiosmotic potential to make ATP during photophosphorylation .
Photosystems
Main article: Photosystem
These photosystems are light-driven redox centers, each consisting of an antenna complex that uses chlorophylls and accessory photosynthetic pigments such as carotenoids and phycobiliproteins to harvest light at a variety of wavelengths. Each antenna complex has between 250 and 400 pigment molecules and the energy they absorb is shuttled by resonance energy transfer to a specialized chlorophyll a at the reaction center of each photosystem.
When either of the two chlorophyll a molecules at the reaction center absorb energy, an electron is excited and transferred to an electron-acceptor molecule.
Chlorophyll and photosynthesis
Chlorophyll is vital for photosynthesis , which allows plants to obtain energy from light.
Chlorophyll molecules are specifically arranged in and around pigment protein complexes
called photosystems which are embedded in the thylakoid membranes of chloroplasts . In these complexes, chlorophyll serves two primary functions. The function of the vast majority of chlorophyll (up to several hundred per photosystem) is to absorb light and transfer that light energy by resonance energy transfer to a specific chlorophyll pair in the reaction center of the photosystems. Because of chlorophyll’s selectivity regarding the wavelength of light it absorbs, areas of a leaf containing the molecule will appear green.
Chemical structure
Chlorophyll is found in high concentrations in chloroplasts of plant cells.
SeaWIFS-derived average sea surface chlorophyll for the period 1998 to 2006.
Chlorophyll is a green pigment found in most plants , algae , and cyanobacteria . Its name is derived from Greek :
χλωρός
( chloros "green") and
φύλλον
( phyllon "leaf"). Chlorophyll absorbs light most strongly in the blue and red but poorly in the green portions of the electromagnetic spectrum , hence the green color of chlorophyll-containing tissues like plant leaves.
[1]
Chlorophyll is vital for photosynthesis , which allows plants to obtain energy from light.
Chlorophyll molecules are specifically arranged in and around pigment protein complexes called photosystems which are embedded in the thylakoid membranes of chloroplasts . In these complexes, chlorophyll serves two primary functions. The function of the vast majority of chlorophyll (up to several hundred per photosystem) is to absorb light and transfer that light energy by resonance energy transfer to a specific chlorophyll pair in the reaction center of the photosystems. Because of chlorophyll’s selectivity regarding the wavelength of light it absorbs, areas of a leaf containing the molecule will appear green.
There are currently two accepted photosystem units, Photosystem II and Photosystem I, which have their own distinct reaction center chlorophylls, named P680 and P700, respectively.
[2] These pigments are named after the wavelength (in nanometers ) of their redpeak absorption maximum. The identity, function and spectral properties of the types of chlorophyll in each photosystem are distinct and determined by each other and the protein structure surrounding them. Once extracted from the protein into a solvent (such as acetone or methanol ), these chlorophyll pigments can be separated in a simple paper chromatography experiment, and, based on the number of polar groups between chlorophyll a and chlorophyll b, will chemically separate out on the paper.
The function of the reaction center chlorophyll is to use the energy absorbed by and transferred to it from the other chlorophyll pigments in the photosystems to undergo a charge separation, a specific redox reaction in which the chlorophyll donates an electron into a series of molecular intermediates called an electron transport chain . The charged reaction center chlorophyll (P680
+
) is then reduced back to its ground state by accepting an electron. In
Photosystem II, the electron which reduces P680
+
ultimately comes from the oxidation of water into O
2
and H + through several intermediates. This reaction is how photosynthetic organisms like plants produce O
2
gas, and is the source for practically all the O
2
in Earth's atmosphere. Photosystem I typically works in series with Photosystem II, thus the P700
+
of
Photosystem I is usually reduced, via many intermediates in the thylakoid membrane, by electrons ultimately from Photosystem II. Electron transfer reactions in the thylakoid membranes are complex, however, and the source of electrons used to reduce P700
+
can vary.
The electron flow produced by the reaction center chlorophyll pigments is used to shuttle H
+ ions across the thylakoid membrane, setting up a chemiosmotic potential mainly used to produce ATP chemical energy, and those electrons ultimately reduce NADP
+
to NADPH a universal reductant used to reduce CO
2
into sugars as well as for other biosynthetic reductions.
Reaction center chlorophyll-protein complexes are capable of directly absorbing light and performing charge separation events without other chlorophyll pigments, but the absorption cross section (the likelihood of absorbing a photon under a given light intensity) is small.
Thus, the remaining chlorophylls in the photosystem and antenna pigment protein complexes associated with the photosystems all cooperatively absorb and funnel light energy to the reaction center. Besides chlorophyll a , there are other pigments, called accessory pigments , which occur in these pigment-protein antenna complexes.
[5]
The different structures of chlorophyll are summarized below:
Chlorophyll a Chlorophyll b
Chlorophyll
c1
Chlorophyll
c2
Chlorophyll d
Molecular formula
C
55
H
72
O
5
N
4
M g
C
55
H
70
O
6
N
4
M g
C
35
H
30
O
5
N
4
M g
C
35
H
28
O
5
N
4
M g
C
54
H
70
O
6
N
4
M g
C3 group -CH=CH
2
-CH=CH
2
-CH=CH
2
-CH=CH
2
-CHO
C7 group -CH
3
-CHO -CH
3
-CH
3
-CH
3
C8 group -CH
2
CH
3
-CH
2
CH
3
-CH
2
CH
3
-CH=CH
2
-CH
2
CH
3
C17 group
-
CH
2
CH
2
COO-
Phytyl
-
CH
2
CH
2
COO-
Phytyl
-
CH=CHCOOH
-
-
CH=CHCOOH
CH
2
CH
2
COO-
Phytyl
C17-C18 bond
Single Single Double Double Single
Occurrenc e
Universal Mostly plants Various algae Various algae cyanobacteria
Structure of chlorophyll a Structure of chlorophyll b
Structure of chlorophyll d
Structure of chlorophyll c1 Structure of chlorophyll c2
Spectrophotometry
Absorbance spectra of free chlorophyll a ( green ) and b ( red ) in a solvent. The spectra of chlorophyll molecules are slightly modified in vivo depending on specific pigment-protein interactions.
Measurement of the absorption of light is complicated by the solvent used to extract it from plant material, which affects the values obtained,
In diethyl ether, chlorophyll a has approximate absorbance
In the electron microscope, thylakoid membranes appear as alternating light-and-dark bands, each 0.01 μm thick. Embedded in the thylakoid membrane is the antenna complex, which consists of the light-absorbing pigments, including chlorophyll and carotenoids , and proteins
(which bind the chlorophyll). This complex both increases the surface area for light capture, and allows capture of photons with a wider range of wavelengths. The energy of the incident photons is absorbed by the pigments and funneled to the reaction centre of this complex through resonance energy transfer . Two chlorophyll molecules are then ionised, producing an excited electron which then passes onto the photochemical reaction centre.
The function of the reaction center chlorophyll is to use the energy absorbed by and transferred to it from the other chlorophyll pigments in the photosystems to undergo a charge separation, a specific redox reaction in which the chlorophyll donates an electron into a series of molecular intermediates called an electron transport chain . The charged reaction center chlorophyll (P680
+
) is then reduced back to its ground state by accepting an electron. In
Photosystem II, the electron which reduces P680
+
ultimately comes from the oxidation of water into O
2
and H
+
through several intermediates. This reaction is how photosynthetic organisms like plants produce O
2
gas, and is the source for practically all the O
2
in Earth's atmosphere.
Photosystem I contains a pair of chlorophyll a molecules, designated P700 , at its reaction center that maximally absorbs 700 nm light. Photosystem II contains P680 chlorophyll that absorbs 680 nm light best (note that these wavelengths correspond to deep red - see the visible spectrum ). The P is short for pigment and the number is the specific absorption peak in nanometers for the chlorophyll molecules in each reaction center.
Cytochrome b6f complex
Main article: Cytochrome b6f complex
The cytochrome b6f complex is part of the thylakoid electron transport chain and couples electron transfer to the pumping of protons into the thylakoid lumen. Energetically, it is situated between the two photosystems and transfers electrons from photosystem IIplastoquinone to plastocyanin-photosystem I.
ATP synthase
Main article: ATP synthase
The thylakoid ATP synthase is a CF1FO-ATP synthase similar to the mitochondrial ATPase.
It is integrated into the thylakoid membrane with the CF1-part sticking into stroma. Thus,
ATP synthesis occurs on the stromal side of the thylakoids where the ATP is needed for the light-independent reactions of photosynthesis.
Thylakoid lumen proteins
The electron transport protein plastocyanin is present in the lumen and shuttles electrons from the cytochrome b6f protein complex to photosystem I. While plastoquinones are lipid-soluble and therefore move within the thylakoid membrane, plastocyanin moves through the thylakoid lumen.
The lumen of the thylakoids is also the site of water oxidation by the oxygen evolving complex associated with the lumenal side of photosystem II.
Lumenal proteins can be predicted computationally based on their targeting signals. In
Arabidopsis, out of the predicted lumenal proteins possessing the "TAT" signal, the largest groups with known functions are 19% involved in protein processing (proteolysis and folding), 18% in photosynthesis, 11% in metabolism, and 7% redox carriers and defense.
[10]
Thylakoid protein expression
Chloroplasts have their own genome , which encodes a number of thylakoid proteins.
However, during the course of plastid evolution from their cyanobacterial endosymbiotic ancestors, extensive gene transfer from the chloroplast genome to the cell nucleus took place.
This results in the four major thylakoid protein complexes being encoded in part by the chloroplast genome and in part by the nuclear genome. Plants have developed several mechanisms to co-regulate the expression of the different subunits encoded in the two different organelles to assure the proper stoichiometry and assembly of these protein complexes. For example, transcription of nuclear genes encoding parts of the photosynthetic apparatus is regulated by light . Biogenesis, stability and turnover of thylakoid protein complexes is regulated by phosphorylation via redox-sensitive kinases in the thylakoid membranes.
[15]
The translation rate of chloroplast-encoded proteins is controlled by the presence or absence of assembly partners (control by epistasy of synthesis).
[16]
This mechanism involves negative feedback through binding of excess protein to the 5' untranslated region of the chloroplast mRNA .
[17]
Chloroplasts also need to balance the ratios of photosystem I and II for the electron transfer chain. The redox state of the electron carrier
plastoquinone in the thylakoid membrane directly affects the transcription of chloroplast genes encoding proteins of the reaction centers of the photosystems, thus counteracting imbalances in the electron transfer chain.
[18]
[
edit
] Thylakoid function
Light-dependent reactions of photosynthesis at the thylakoid membrane
The thylakoids are the site of the light-dependent reactions of photosynthesis. These include light-driven water oxidation and oxygen evolution , the pumping of protons across the thylakoid membranes coupled with the electron transport chain of the photosystems and cytochrome b6f complex, and ATP synthesis by the ATP synthase utilizing the generated proton gradient.
Water photolysis
The first step in photosynthesis is the light-driven oxidation (splitting) of water to provide the electrons for the photosynthetic electron transport chains as well as protons for the establishment of a proton gradient. The water-splitting reaction occurs on the lumenal side of the thylakoid membrane and is driven by the light energy captured by the photosystems. It is interesting to note that this oxidation of water conveniently produces the waste product O
2 that is vital for cellular respiration . The molecular oxygen formed by the reaction is released into the atmosphere.
Electron transport chains
Two different variations of electron transport are used during photosynthesis:
Noncyclic electron transport or Non-cyclic photophosphorylation produces
NADPH + H
+
and ATP.
Cyclic electron transport or Cyclic photophosphorylation produces only ATP.
The noncyclic variety involves the participation of both photosystems, while the cyclic electron flow is dependent on only photosystem I.
Photosystem I uses light energy to reduce NADP
+
to NADPH + H
+
, and is active in both noncyclic and cyclic electron transport. In cyclic mode, the energized electron is passed down a chain that ultimately returns it (in its base state) to the chlorophyll that energized it.
Photosystem II uses light energy to oxidize water molecules, producing electrons (e -
), protons (H
+
), and molecular oxygen (O
2
), and is only active in noncyclic transport.
Electrons in this system are not conserved, but are rather continually entering from oxidized 2H
2
O (O
2
+ 4 H
+
+ 4 e
-
) and exiting with NADP
+
when it is finally reduced to NADPH.
Chemiosmosis
A major function of the thylakoid membrane and its integral photosystems is the establishment of chemiosmotic potential. The carriers in the electron transport chain use some of the electron's energy to actively transport protons from the stroma to the lumen .
During photosynthesis, the lumen becomes acidic , as low as pH 4, compared to pH 8 in the stroma . This represents a 10,000 fold concentration gradient for protons across the thylakoid membrane.
Source of proton gradient
The protons in the lumen come from three primary sources.
Photolysis by photosystem II oxidises water to oxygen , protons and electrons in the lumen.
The transfer of electrons from photosystem II to plastoquinone during non-cyclic electron transport consumes two protons from the stroma. These are released in the lumen when the reduced plastoquinol is oxidized by the cytochrome b6f protein complex on the lumen side of the thylakoid membrane. From the plastoquinone pool, electrons pass through the cytochrome b6f complex. This integral membrane assembly resembles cytochrome bc1.
The reduction of plastoquinone by ferredoxin during cyclic electron transport also transfers two protons from the stroma to the lumen.
The proton gradient is also caused by the consumption of protons in the stroma to make
NADPH from NADP+ at the NADP reductase.
ATP generation
The molecular mechanism of ATP generation in chloroplasts is similar to that in mitochondria and takes the required energy from the proton motive force (PMF). However, chloroplasts rely more on the chemical potential of the PMF to generate the potential energy required for ATP synthesis. The PMF is the sum of a proton chemical potential (given by the proton concentration gradient) and a transmembrane electrical potential (given by charge separation across the membrane). Compared to the inner membranes of mitochondria, which have a significantly higher membrane potential due to charge separation, thylakoid membranes lack a charge gradient. To compensate for this, the 10,000 fold proton concentration gradient across the thylakoid membrane is much higher compared to a 10 fold gradient across the inner membrane of mitochondria. The resulting chemiosmotic potential between the lumen and stroma is high enough to drive ATP synthesis using the ATP synthase . As the protons travel back down the gradient through channels in ATP synthase ,
ADP + P i
is combined into ATP. In this manner, the light-dependent reactions are coupled to the synthesis of ATP via the proton gradient.
Recent studies have shown that chloroplasts can be interconnected by tubular bridges called stromules , formed as extensions of their outer membranes.
[6][7]
Chloroplasts appear to be able to exchange proteins via stromules,
[8]
and thus function as a network. [ edit ] Chlorophyll and photosynthesis
Chlorophyll is vital for photosynthesis , which allows plants to obtain energy from light.
Chlorophyll molecules are specifically arranged in and around pigment protein complexes called photosystems which are embedded in the thylakoid membranes of chloroplasts . In these complexes, chlorophyll serves two primary functions. The function of the vast majority of chlorophyll (up to several hundred per photosystem) is to absorb light and transfer that light energy by resonance energy transfer to a specific chlorophyll pair in the reaction center of the photosystems. Because of chlorophyll’s selectivity regarding the wavelength of light it absorbs, areas of a leaf containing the molecule will appear green.
There are currently two accepted photosystem units, Photosystem II and Photosystem I, which have their own distinct reaction center chlorophylls, named P680 and P700, respectively.
[2]
These pigments are named after the wavelength (in nanometers ) of their redpeak absorption maximum. The identity, function and spectral properties of the types of chlorophyll in each photosystem are distinct and determined by each other and the protein structure surrounding them. Once extracted from the protein into a solvent (such as acetone or methanol ), these chlorophyll pigments can be separated in a simple paper chromatography experiment, and, based on the number of polar groups between chlorophyll a and chlorophyll b, will chemically separate out on the paper.
The function of the reaction center chlorophyll is to use the energy absorbed by and transferred to it from the other chlorophyll pigments in the photosystems to undergo a charge separation, a specific redox reaction in which the chlorophyll donates an electron into a series of molecular intermediates called an electron transport chain . The charged reaction center chlorophyll (P680
+
) is then reduced back to its ground state by accepting an electron. In
Photosystem II, the electron which reduces P680 + ultimately comes from the oxidation of water into O
2
and H
+
through several intermediates. This reaction is how photosynthetic organisms like plants produce O
2
gas, and is the source for practically all the O
2
in Earth's atmosphere. Photosystem I typically works in series with Photosystem II, thus the P700
+
of
Photosystem I is usually reduced, via many intermediates in the thylakoid membrane, by electrons ultimately from Photosystem II. Electron transfer reactions in the thylakoid membranes are complex, however, and the source of electrons used to reduce P700
+
can vary.
The electron flow produced by the reaction center chlorophyll pigments is used to shuttle H + ions across the thylakoid membrane, setting up a chemiosmotic potential mainly used to produce ATP chemical energy, and those electrons ultimately reduce NADP
+
to NADPH a universal reductant used to reduce CO
2
into sugars as well as for other biosynthetic reductions.
Reaction center chlorophyll-protein complexes are capable of directly absorbing light and performing charge separation events without other chlorophyll pigments, but the absorption cross section (the likelihood of absorbing a photon under a given light intensity) is small.
Thus, the remaining chlorophylls in the photosystem and antenna pigment protein complexes
associated with the photosystems all cooperatively absorb and funnel light energy to the reaction center. Besides chlorophyll a , there are other pigments, called accessory pigments , which occur in these pigment-protein antenna complexes.
Information provided by: http://gened.emc.maricopa.edu
Photosynthesis: Pathway of
Carbon Fixation
Photosynthesis is the synthesis of organic molecules using the energy of light. For the sugar glucose (one of the most abundant products of photosynthesis) the equation is:
6CO
2
+ 12H
2
O -> C
6
H
12
O
6
+ 6H
2
O + 6O
2
Light provides the energy to:
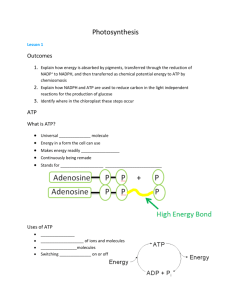
transfer electrons from water to nicotinamide adenine dinucleotide phosphate ( NADP + ) forming NADPH ;
generate ATP .
The details of these processes are described in Photosynthesis: The Role of Light .
ATP and NADPH provide the energy and electrons to reduce carbon dioxide
(CO
2
) to organic molecules.
The Steps
CO
2
combines with the phosphorylated 5-carbon sugar ribulose bisphosphate .
This reaction is catalyzed by the enzyme ribulose bisphosphate
carboxylase oxygenase ( RUBISCO - an enzyme which can fairly claim to be the most abundant protein on earth).
The resulting 6-carbon compound breaks down into two molecules of 3phosphoglyceric acid ( PGA ).
The PGA molecules are further phosphorylated (by ATP ) and are reduced
(by NADPH ) to form phosphoglyceraldehyde ( PGAL ).
Phosphoglyceraldehyde serves as the starting material for the synthesis of
glucose and fructose .
Glucose and fructose make the disaccharide sucrose , which travels in
solution to other parts of the plant (e.g., fruit, roots).
Glucose is also the monomer used in the synthesis of the polysaccharides starch and cellulose .
The graphic shows the steps in the fixation of carbon dioxide during photosynthesis. These steps were worked out by Melvin Calvin and his colleagues at the University of California and, for this reason, are named the Calvin cycle.
Link to graphic showing how they did it.
The first step involves the addition of CO
2
to a 5-Carbon acceptor
[RUBP].
This leads to the production of two 3-Carbon molecules.
The next step involves the reduction of the 3-Carbon molecule into a Carbohydrate .
The final step is a complex series of reactions, which regenerates the
5-Carbon Acceptor
(Ribulose-1-5-bisphosphate [ RUBP ]).
One CO 2 is gained with each cycle . Six cycles lead to the formation of a 6-Carbon sugar.
Calvin cycle
The series of dark reactions, also known as the carbon fixation stage, which forms the second of the two stages of photosynthesis after photolysis . The Calvin cycle takes place in the stroma of chloroplasts .
Carbon dioxide enters the cycle, together with energy made available by photolysis, and in a series of steps, glucose is manufactured. This glucose can be used in respiration or as a building block in making starch and cellulose .
Related category
•
BIOCHEMISTRY
Also on this site:
Encyclopedia of Alternative Energy & Sustainable Living
Encyclopedia of History
Transport Concepts & Designs (partner site)
BACK TO TOP