Endocrine control of stallion reproduction

Kufa University
Veterinary Medicine college
Theriogenology Fertility Majid .A .Talal
HORMON CONTROL OF The male
Reproductive System
1
2
Endocrine control of reproduction;-
Gonadotrophin release is regulated primarily by the episodic stimulation of the pituitary gland by gonadotrophin releasing hormone (GnRH)
• Luteinising hormone (LH) and, to a lesser extent, follicle stimulating hormone (FSH), are secreted into the peripheral circulation in an episodic manner by the anterior pituitary gland.
• The functional significance of this pulsatile release is not known.
• LH secretion is immediate but transitory after GnRH stimulation.
• FSH secretion is slow and gradual
.
• The half-life of FSH is also longer than that of LH, therefore the plasma changes in LH are more rapid than those of FSH.
• LH binds to specific receptors on Leydig cells of the testis and stimulates testicular steroidogenesis.
• Testosterone secretion occurs both locally within the testis and into the peripheral circulation.
• Peripheral concentrations of testosterone are necessary for the development and maintenance of secondary sexual characteristics, maintenance of sexual behaviour and negative feedback regulation of gonadotrophin secretion. The production of testosterone within the testis is important for initiating and maintaining spermatogenesis.
• High intratesticular testosterone concentrations are maintained within the seminiferous tubules, in part by the binding of testosterone to androgen binding protein.
• FSH binds to specific receptors on the plasma membrane of
Sertoli cells.
3
• Androgen binding protein is produced by the Sertoli cells under the influence of testosterone and FSH.
• FSH binding results in the production of a variety of proteins that are important in regulating spermatogenesis.
• Feedback control of FSH is via the gonadal peptide inhibin .
• Inhibin is a non-steroidal glycoprotein product of the Sertoli cells that selectively inhibits FSH secretion at the pituitary gland.
• Other negative feedback mechanisms are important: testosterone and its active metabolites oestradiol and dihydrotestosterone exert a profound suppressive effect upon both LH and FSH secretion.
• The Sertoli cells also produce other substances, termed activins , which have a stimulatory effect upon pituitary FSH secretion.
• Oestrogens probably have similar functions to androgens, but it is not known why the stallion produces such large quantities of these hormones, i.e. 17 b
-oestradiol sulphate and oestrone sulphate.
• Circulating blood concentrations of testosterone and oestrogens vary considerably from hour to hour, so that measurement of a single sample is often meaningless; concentrations particularly of oestrogens are usuallyhighest during the breeding season.
• Stallions will usually copulate all year round, but most reproductive parameters are maximal during the breeding season (April to
September)
• Inactive stallions may have lower mean testosterone concentrations thanthose that are mating.
• Exposure of a stallion to a mare in oestrus causes a rise in circulating oestrone sulphate concentrations within ten minutes.
4
• The reproductive season can be brought forward, as in the mare, by increasing artificial light; in some stallions this also produces a
‘ premature autumn ’ .
• Puberty, as judged by the time when the stallion ’ s ejaculate contains a minimum of 100 \ 106 spermatozoa with at least 10% progressive motility, occurs at about 18 months; this may be influenced by the time of birth,nutritional status and breed and can be considerably delayed in certain
Individuals
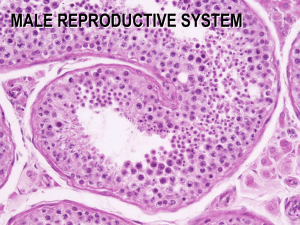
Spermatogenesis, or the process of sperm (spermatozoon) production, is composed of two processes: spermatocytogenesis and spermiogenesis. Spermatocytogenesis ; is the development of spermatogenic tissue in the testis, which occurs as a result of cell division of the spermatogonia, located at the periphery of the testicular lobules (see Fig. 2.6). As the cells divide they move progressivelytowards the lumen of the lobule. The spermatogonia undergo several mitotic cell divisions (where the number of chromosomes remain the same), eventually forming the large primary spermatocytes.
A meiotic or reduction cell division then occurs in which secondary spermatocytes are formed. In this meiotic division the number of chromosomes is reduced from the normal or diploid (60 chromosomes) to the haploid (30 chromosomes) state. This reduction division is analogous to that which occurs in the female primary oocyte (see Chapter 4).
5
After the meiotic division of the primary spermatocytes, the secondary spermatocytes divide further to form smaller cells, the spermatids (Fig. 3.3).
Eachsecondary spermatocyte produces two spermatids. During the
6
subsequentphase, spermiogenesis, the spermatids undergo differentiation, each one eventually forming a sperm. The changes include formation of the acrosome ornuclear cap, the head, midpiece and tail
In the process a droplet of the spermatid cytoplasm is retained temporarilybetween the head and midpiece of the sperm and this is often seen in immaturespermatozoa. The whole process of spermatogenesis takes approximately60 days in the bull. Each sperm cell weighs approximately 2.5 ¥ 10-11 g, half ofwhich is contained in the head. The head is largely taken up by the nucleus,which contains the genetic material.
Spermatogenesis is essentially controlled by follicle stimulating hormone(FSH), which appears to act on the Sertoli cells. These in turn control the rateof spermatogenesis, although the mechanism for this is unclear. The presenceof adequate concentrations of testosterone is also required.The Sertoli cellsare also the source of inhibin, a polypeptide hormone which enters the systemiccirculation to selectively suppress FSH secretion by the pituitary gland. The hormonal relationships controlling testosterone production and spermatogenesisare illustrated in Fig.
3.5.
Hormone secretion in the male is pulsatile, as in the female. Each pulse ofLH and FSH is followed by a pulse of testosterone (see
Fig. 3.1). Testosteronein turn exerts a negative feedback effect, suppressing further pulses of LH secretion for a variable period of time.
7