Chapter 3
The Chromosomal Basis of
Heredity
Chromosomes
• The chromosome complement = the complete set
of chromosomes of plants and animals
• The nucleus of each somatic cell contains a fixed
number of chromosomes typical of the particular
species
• The number of chromosomes vary tremendously
among species and have little relationship to the
complexity of the organism
2
Chromosomes
• The chromosomes in the nuclei of somatic cells
are usually present in pairs. For example, the 46
chromosomes of human being consist of 23
pairs
• Cells with nuclei of this sort, containing two
similar sets of chromosomes, are called diploid
3
Chromosomes
• The germ cells, or gametes, are haploid and
contain only one set of chromosomes, consisting
of one member of each of the pairs
• The haploid gametes unite in fertilization to
produce the diploid state of somatic cell
• The chromosomes are present in pairs because
one chromosome of each pair derives from the
maternal parent of the organism and the other
from its paternal parent
4
Mitosis
• Mitosis is a precise process of nuclear division that
ensures that each of two daughter cells receives a
diploid complement of chromosomes identical with
the diploid complement of the parent cell
• Mitosis is usually accompanied by cytokinesis, the
process in which the cell itself divides to yield two
daughter cells
5
Cell Cycle
• In a cell that is not undergoing mitosis, the
chromosomes are invisible with a light microscope.
This stage of the cell cycle is called interphase
• DNA in the chromosomes is replicated during a
period of interphase called S = DNA synthesis
• Before and after S, there are periods, called G1 and
G2, respectively
• These three interphase periods are followed by
mitosis, M
6
Figure 3.2: The cell cycle of a typical mammalian cell growing in tissue
culture
7
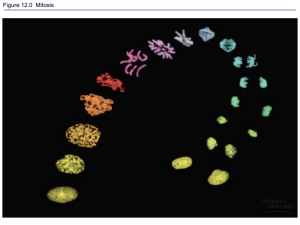
Stages of Mitosis
• Prophase is marked by the condensation of
chromosomes. Each chromosome is already
longitudinally double, consisting of two subunits
called chromatids
• Each pair of chromatids is the product of the
duplication of one chromosome in the S period of
interphase
• The chromatids in a pair are held together at a
specific region of the chromosome called the
centromere.
8
Prophase of Haemanthus
© Andrew S. Bajer - Research Projects
9
Stages of Mitosis
• At the beginning of metaphase, the mitotic spindle
forms
• The spindle is a bipolar structure arching between
the centrosomes that consists of microtubules
• The spindle fibers attach to each chromosome in
the region of the centromere at the kinetochore
• The chromosomes move toward the center of the
cell until all the kinetochores lie on an imaginary
plane equidistant from the spindle poles = the
metaphase plate
10
Metaphase of Haemanthus
© Andrew S. Bajer - Research Projects
11
Stages of Mitosis
• In anaphase, the centromeres divide
longitudinally, and the two sister chromatids of
each chromosome move toward opposite poles
of the spindle
• Once the centromeres divide, each sister
chromatid is regarded as a separate
chromosome in its own right.
12
Anaphase of Haemanthus
© Andrew S. Bajer - Research Projects
13
Stages of Mitosis
• In telophase, a nuclear envelope forms around
each compact group of chromosomes, nucleoli
are formed, and the spindle disappears
• The chromosomes undergo decondensation until
they are no longer visible as discrete entities
• The two daughter nuclei assume a typical
interphase appearance
• The cytoplasm of the cell divides in two
14
Telophase of Haemanthus
© Andrew S. Bajer - Research Projects
15
Figure 3.3: Chromosome behavior during mitosis in an organism with two
16
pairs of chromosomes
Meiosis
• Meiosis is a mode of cell division in which cells are
created that contain only one member of each pair of
chromosomes
• Meiosis consists of two successive nuclear divisions
• Meiosis results in four daughter cells, each genetically
different and each containing one haploid set of
chromosomes
• Meiosis is a more complex and considerably longer
process than mitosis and usually requires days or
even weeks
17
Meiosis
• In animals, meiosis takes place in specific cells
called meiocytes
• The oocytes form egg cells and the spermatocytes
form sperm cells
• In the females of both animals and plants, only one
of the four products develops into a functional cell
(the other three disintegrate)
18
Figure 3.5: The life cycle of a typical animal
19
Meiosis
• In plants, the products of meiosis form spores,
which undergo one or more mitotic divisions to
produce a haploid gametophyte organism
• The gametophyte produces gametes by mitotic
division of a haploid nucleus
• Fusion of haploid gametes creates a diploid zygote
that develops into the sporophyte plant, which
undergoes meiosis to produce spores and so
restarts the cycle
20
Figure 3.6: The life cycle of corn, Zea mays
21
Outline of Meiosis
• Prior to the first nuclear division, the members of
each pair of chromosomes become closely
associated along their length
• The chromosomes that pair with each other are
said to be homologous chromosomes
• Each member of a pair of homologs consists of a
duplex of two sister chromatids joined at the
centromere. The pairing of the homologous
chromosomes, therefore, produces a fourstranded structure
22
Outline of Meiosis
• At the time of pairing, the homologs can exchange
genes that results in chromosomes that consist of
segments from one homolog intermixed with
segments from the other
• In the first nuclear division, the homologous
chromosomes are separated from each other, one
member of each pair going to opposite poles of the
spindle
• Two nuclei are formed, each containing a haploid
set of duplex chromosomes
23
Outline of Meiosis
• The second nuclear division resembles a
mitotic division, but there is no DNA
replication
• At metaphase, the chromosomes align on
the metaphase plate, and at anaphase,
the chromatids are separated into
opposite daughter nuclei
• The net effect of the two divisions is the
creation of four haploid nuclei, each
containing the equivalent of a single
sister chromatid from each pair of
homologous chromosomes
Figure 3.4: Behavior of a single pair of homologous chromosomes in meiosis
24
Mitosis vs. Meiosis
• Meiosis produces four cells: each contains
one copy of each pair of homologous
chromosomes = genetically different,
haploid
• Mitosis produces two cells that contain both
members of each pair of homologous
chromosomes = genetically identical, diploid
25
Meiosis I
• The first meiotic division—reductional division,
reduces the chromosome number by half
• Prophase I is the longest stage and is commonly
divided into five substages: leptotene, zygotene,
pachytene, diplotene, and diakinesis
These are descriptive terms that indicate the
appearance of the chromosomes at each substage
26
Meiosis: Prophase I
• Leptotene – the chromosomes first
become visible as long, thread-like
structures
Figure 3.8A: Leptotene
• Zygotene – synapsis of homologous
chromosomes = bivalent
Figure 3.8B: Zygotene
• Pachytene – crossing-over between homologs
Figure 3.8C: Early
pachytene
Parts A, B, and C courtesy of Marta Walters and Santa Barbara Botanic
Gardens, Santa Barbara, California. Part D courtesy of Herbert Stern.
Used with permission.
Figure 3.8D: Late
pachytene
27
Figure 3.9: Bivalent consisting of a pair of homologous chromosomes
28
Meiosis: Prophase I
• Diplotene – chromosome repulsion, however, they remain held
together by cross-connections resulting from crossing-over.
Each cross-connection, called a chiasma, is formed by a
breakage and rejoining between nonsister chromatids
• Diakinesis – maximum chromosome contraction
29
Meiosis: Metaphase I
• Metaphase I – the bivalents positioned with the
centromeres of the two homologs on opposite
sides of the metaphase plate
• As each bivalent moves onto the
metaphase plate, its centromeres
are oriented at random with respect
to the poles of the spindle
• Genes on different chromosomes undergo independent
assortment because nonhomologous chromosomes align
at random in metaphase I
30
Figure 3.11: Independent assortment of genes on nonhomologous
chromosomes
31
Meiosis: Anaphase I
• Anaphase I – homologous
chromosomes, each composed of
two chromatids joined at an
undivided centromere, separate
from one another and move to
opposite poles of the spindle
• The physical separation of homologous
chromosomes in anaphase I is the physical basis of
Mendel’s principle of segregation
32
Meiosis: Telophase I
• Telophase I – a haploid set of
chromosomes consisting of one
homolog from each bivalent is
located near each pole of the spindle
• The spindle breaks down, the
chromosomes enter the second
meiotic division after only a limited
uncoiling
• Chromosome replication never takes place
between the two divisions
33
Meiosis II
• The second meiotic division (meiosis II) is called the
equational division because the chromosome
number remains the same in each cell before and
after the second division
• In some species, the chromosomes pass directly
from telophase I to prophase II without loss of
condensation
• After a short prophase II and the formation of
second-division spindles, the centromeres of the
chromosomes in each nucleus become aligned on
the central plane of the spindle at metaphase II
34
Meiosis II
• In anaphase II, the centromeres divide and the
chromatids of each chromosome move to opposite
poles of the spindle
• Once the centromere has split at anaphase II, each
chromatid is considered a separate chromosome
• Telophase II is a transition to the interphase
condition of the chromosomes in the four haploid
nuclei, accompanied by division of the cytoplasm.
35
Meiosis
• The chromatids of a chromosome are
usually not genetically identical because of
crossing-over associated with the
formation of chiasmata during prophase of
the first division
36
Figure 3.7: Chromosome behavior during meiosis
37
Chromosome Structure
• Eukaryotic chromosomes are highly coiled stable
complexes of DNA and protein called chromatin
• Each eukaryotic chromosome contains a single DNA
molecule of enormous length
• Some of the proteins present in chromatin determine
chromosome structure and the changes in structure
during the cell cycle
• Other chromatin proteins appear to have important
roles in regulating chromosome functions
38
Chromatin Structure
• The nucleosome is the basic structural unit of
chromatin
• Each nucleosome is composed of a core particle,
~55 base pairs of DNA called linker DNA that links
adjacent core particles and one molecule of histone
H1 that binds to the core particle and to the linker
DNA
• Histones are small proteins that are highly
conserved among different organisms
39
Chromatin Structure
• Each core particle consists of an octamere of
pairs each of histone H2A, H2B, H3, and H4; a
segment of DNA containing about 145 base pairs
Figure 3.15a: Organization of nucleosomes
40
Figure 3.15b: Organization of nucleosomes
41
Chromatin Structure
• In the nucleus of a nondividing
cell, chromatin fibers form discrete
chromosome territories
• Chromosome territories are
correlated with gene densities
• Territories of chromosome
domains that are relatively gene
rich tend to be located toward the
interior of the nucleus
Figure courtesy of Tobias A. Knoch, Erasmus MC, Rotterdam, and
Kirchhoff-Institute for Physics, Ruperto-Carola University, Heidelberg
Figure 3.18: Chromosome
territories formed by 30nm chromatin fibers within
the nucleus of a
nondividing cell
42
• Nucleosomes coil to
form higher order DNA
structure called the 30nm chromatin fiber
Figure 3.19: Condensation of DNA and
chromatin to form a metaphase
chromosome
43
Chromatin Structure
• The spaces between the chromatin domains form a
network of channels large enough to allow passage
of the molecular machinery for replication,
transcription, and RNA processing
• Replication takes place in small discrete regions that
exhibit a reproducible temporal and spatial pattern,
and transcription takes place in a few hundred
discrete locations
• The metaphase chromosome is a hierarchy of coiled
coils
44
Chromatin Structure
• Compact and heavily stained regions of chromatin
are known as heterochromatin, which mainly
consists of highly repeated noncoding DNA
sequences—satellite DNA
• The rest of the chromatin, which becomes visible
only after chromosome condensation in mitosis or
meiosis, is called euchromatin
• The number of genes located in heterochromatin
is small relative to the number in euchromatin
45
Figure 3.21a: Metaphase
chromosomes of the ground
squirrel
Figure 3.21B: An interpretive drawing of
metaphase chromosomes of the ground
squirrel
Part A courtesy of T.C. Hsu, Ph.D., and used with permission of Sen
Pathak, Ph.D., Anderson Cancer Center, University of Texas.
46
Chromosome Structure
• The centromere is essential for chromosome
segregation
• The centromere is a specific region of the
eukaryotic chromosome. It serves as a central
component of the kinetochore the complex of
DNA and proteins to which the spindle fibers
attach and move the chromosomes in both
mitosis and meiosis
47
Figure 3.22: A yeast centromere
Adapted from K. S. Bloom, M. Fitzgerald-Hayes, and J. Carbon, Cold
Spring Harb. Symp. Quant. Biol. 47 (1982): 1175.
48
Chromosome Structure
• The telomere is essential for the stability of the
chromosome tips
• Due to the nature of DNA replication, chromosomes
require special mechanism to restore DNA in
telomeres in each cycle of replication
• The mechanism relies on an enzyme called telomerase
49
Figure 3.25: The function of telomerase
50
Figure 3.26: Telomere formation in Tetrahymena
51
Chromosomes and Heredity
• Chromosome Theory of Heredity: Genes are
located in chromosomes
• Early evidence that genes are located on
chromosomes was found by Thomas Hunt Morgan
in 1910
• Morgan’s studied inheritance patterns in
Drosophila melanogaster and found that in some
cases reciprocal crosses yield different results
52
Morgan’s Fruit Fly Experiments
• Morgan realized that it might happen if the alleles
for some genes were present in the X
chromosome
• The X chromosome is transmitted in a different
pattern by males and females, and the Y
chromosome does not contain alleles
homologous to genes on the X chromosome
53
Figure 3.29: A chromosomal interpretation of results obtained in F1 and F2
progenies in crosses of Drosophila
54
Nondisjunction
• Experimental proof of the chromosome theory of
heredity came from nondisjunction
• Nondisjunction = chromosomes fail to separate
(disjoin) and move to opposite poles of the division
spindle, results in loss or gain of a chromosome
• Calvin Bridges demonstrated that exceptional
behavior of chromosomes is precisely paralleled
by exceptional inheritance of their genes
55
Figure 3.32: The results of meiotic nondisjunction of the X chromosomes in
a female Drosophila
56
X-Linked Inheritance
• Special chromosomes determine sex in many organisms
• X and Y chromosomes = sex chromosomes, which are nonidentical but share some genes
• In most organisms, the Y chromosome carries few genes other
than those related to male determination
• X-linked genes are inherited according to sex
• Hemophilia is a classic example of human X-linked inheritance
57
X-Linked Inheritance
• In many organisms, the male is the heterogametic
sex
• Males produce two different types of gametes: one
containing X and another Y chromosome
• Females have two X chromosomes and produce
only X-bearing gametes
• In some organisms (birds, butterflies, and some
reptiles), females are heterogametic
58
Data analysis
• Genetic data analysis makes use of probability and
statistics
• Progeny of crosses are predicted by the binomial
probability
• If the probability of possibility A is p and the
probability of the alternative possibility B is q, then
the probability that, in n trials, A is realized s times
and B is realized t times is
n! psqt
s!t!
59
Chi-Square Analysis
• The test of goodness of fit = test analyzes whether
observed data agree with theoretical expectation
• A conventional measure of goodness of fit is a
value called chi-square, c2
• c2 = ∑(observed – expected)2 / expected
•
A value of c2 = 0 means that the observed
numbers fit the expected numbers perfectly
60
Chi-Square Analysis
• Probability P that a worse fit (or one equally bad)
would be obtained by chance, assuming that the
genetic hypothesis is true
• The critical values of P are conventionally chosen
as 0.05 (the 5 percent level) and 0.01 (the 1 percent
level)
• Statistically significant refers to the magnitude of
the difference between the observed and the
expected numbers
61
Chi-Square Analysis
• To determine the P value corresponding to a
calculated c2 we need the number of degrees of
freedom of the particular chi-square test
• The number of degrees of freedom equals the
number of classes of data minus 1
62
Figure 3.34: Graphs for interpreting goodness of fit to genetic predictions
using the chi-square test
63