PowerPoint 簡報
advertisement

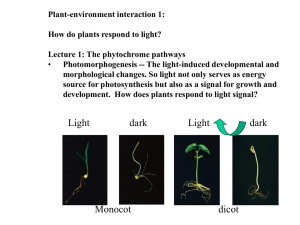
17 Photomorphogenesis: responding to light Fig. 9 18 Tab. 1 2 3 4 5 6 7 8 10 11 12 13 14 15 16 17 1 2 3 4 Light perception in plants • Because plants do not enjoy the luxury of being able to change their environment or seek shelter from adverse conditions by changing their location, they must be more sensitive to changes in their surrounding so they can adapt accordingly. • Plants can sense light gradients and detect subtle differences in spectral composition. Photomorphogenesis • Photomorphogenesis is referring to the response of plant to light, which is the central theme in plant development. Photoreceptors • Most photomorphogenic responses in higher plants appear to be under control of one (or more) of four classes of photoreceptors: 1. Phytochromes (red and far-red) 2. Cryptochrome (blue and UV-A): seedling development and flowering 3. Phototropin (blue and UV-A): differential growth in a light gradient 4. UV-B receptors: unknown Chapter outline • Red and far-red responses • Blue and UV-A responses • Interactions between photoreceptorsUV-B responses Phytochromes • Phytochromes are plants photoreceptors. • Phytochromes are photochromic. They can absorb red (665nm) and far-red (730nm) light and they have two forms, redabsorbing form (Pr) and far red-absorbing form (Pfr). Figure 17.1 Phytochrome is photoreversible • Pr and Pfr forms of phytochrome can change to the other form when expose to red or far-red light, respectively. Phytochrome is photoreversible The photoreversibility of phytochrome comes from its chromophore, phytochromobilin (PΦB) PΦB is covalently linked with the N-terminal part of phytochrome Conformational change of PΦB results in Pr Pfr change Pfr form is the active from of phytochrome Phytochrome is down regulated after activation • The down regulation of phytochrome involved mRNA and protein degradation. • Also, the expression of phytochrome will be down regulated at transcriptional level after activation. Figure 17.5 declines because Pfr is declining. is relatively unstable, with a half life (t1/2) of 1~1.5hr Figure 17.7 Five seconds of red light causes mRNA level declines 15 minutes of lag period follows mRNA drops 50% within the first hour mRNA drops 95% within first two hours Pr and Pfr forms of phytochrome is always in a dynamic equilibrium Phytochrome responses can be grouped into three groups 1~1000 mmol/m2 10-6~10-3 mmol/m2 Very Low Fluence Responses (VLFRs) 0.1nmol/m2 ~ 50 nmol/m2 only converts less than 0.01% of total phytochrome to Pfr form Because far-red light can only convert 97% Pfr to Pr, which is more than what needed to induce VLFRs, so VLFRs are not reversible Very Low Fluence Responses (VLFRs) the principle evidence that VLFRs is mediated by phytochrome is the similarity of its action spectrum to the absorption spectrum of Pr. Most VLFRs are related to germination. It obeys the law of reciprocity. It peaks at red and blue. Low Fluence Responses (LFRs) 1~1000 mmol/m2 Seed germination Seedling development Bioelectric potentials and ion distribution Photoreversible Exhibit reciprocity between duration of irradiation and fluence rate Peaks at red and far-red LFR is induced by poising the system with a maximum level of Pfr for a very brief period of time. LFRs in seed germination positively photoblastic – germination stimulated by light negatively photoblastic – germination inhibited by light A one mm thickness of fine soil will block more than 99% of light. Only light with wavelength longer than 700nm will be able to pass. Very little Pfr is required to stimulate germination. LFRs in seedling development de-etiolation of seedlings Figure 18.10 Table 17.3 LFRs in bioelectric potentials and ion distribution phytochrome-induced changes in the surface potential of the dark-grown barley roots (T. Tanada) red light root tip become positively charged far-red light root tip restore its negative charge Red light induces a depolarization of the membrane within 5-10s following a red light treatment. Subsequent far-red treatment causes a slow return to normal polarity or small hyperpolarization. Figure 18.11 Nyctinastic (sleep) movement Pulvinus (bulbous zone) at the base of leaf/leaflet will drive leaf movement by altering its shape as a result of differential changes in the volume of cells on the upper and lower side of the organ. Pulvinus is osmotically driven by rapid redistribution of K+, Cl- and malate. Pulvinus is osmotically driven by rapid redistribution of K+, Cl- and malate. H+ efflux K+ channels open High Irradiance Responses (HIRs) -prolonged/continuous exposure to light (far-red or direct sunlight) of relatively high irradiance -Response is proportional to the irradiance within a certain range (That’s why they are called HIRs, not HFRs.) -Not photoreversible -Not obeying the law of reciprocity -Many of them are also LFRs Example 1: anthocyanin synthesis Example 2: Inhibition of stem elongation Example 1: Anthocyanin synthesis The initiation of anthocyanin accumulation is classical LFR, peaks at red region. However, when the duration of irradiation lengthens, peak shifts from R FR. Example 2: Inhibition of stem elongation in white mustard Only dark-grown tissue respond to far-red. Green tissue is more responsive to red light. During de-etiolation, HIR peak shifts from far-red to red. Light-grown Dark-grown Phytochrome under natural conditions • Under natural conditions, phyA may just detect the presence/absence of light since it only accumulate under dark-grown conditions. • Other phytochrome response observed under natural conditions is shade avoidance syndrome. Light under canopy is far-red enriched FR+R Far-red Shade avoidance is triggered by far-red light, which can be shown in end-of-day treatment Shade-avoidance syndrome Figure 17.14 Phytochrome signal transduction • Phytochrome is a protein kinase. • When activated, it will phosphorylate other proteins and begin signal pathways. Proteins that are phosphorylated by phytochrome Phytochrome regulates gene expression • A lot of nuclear-encoded genes are regulated by phytochromes, including the small subunit of rubsico (RBCS) and the light-harvesting chlorophyll a/b binding proteins (CAB). • Some proteins are positively regulated, like RBCS and CAB; others are negatively regulated, like phyA and NADPH-protochlorophyllide oxidoreductase. Figure 17.17 Phytochrome also regulates other transcription factor’s activities • PIF3 (phytochrome interacting factor 3) is a transcription activator. • When phytochrome activates (Pr Pfr), the Pfr form binds to PIF3 and activates it. Then activated PIF3 will activate transcription of a large variety of proteins containing G-box motifs. Figure 17.18 Blue and UV-A light responses Cryptochrome Phototropin Cryptochrome is a flavoprotein 5,10-methenyltetrahydrofolate Cryptochromes • Cryptochromes are blue/UV-A photoreceptors mediating seedling development/flowering responses in plants. • In Arabidopsis, there are two cryptochromes, cry1 and cry2. The structure of cry2 is also similar to cry1 with two chromophores. • Cry2 has a role in determining flowering time. Phototropin • Phototropin was orginally isolated as nph1 (nonphototropic hypocotyl 1). Phototropin • Phototropin is also a flavoprotein with two flavin mononucleotide (FMN) chromophores. • FMN chromophores binds to domain called LOV (light, oxygen and voltage) domain. Phototropin could be a blue-light dependent protein kinase Interactions between Photoreceptors 100% 20% 68% Hook straightening and cotyledon unfolding are controlled by all three photoreceptors Cotyledon expansion is controlled by phyB and cry1 phyB controls hypocotyl elongation CAB genes can be induced by either phyA (VLFR) or phyB (LFR).