20140916-003329
advertisement

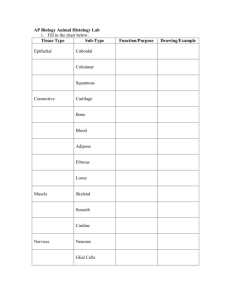
The bone The cartilage Muscle tissue Dr. Makarchuk Iryna Bone is a connective tissue characterized by a mineralized extracellular matrix. • Bone is a specialized form of connective tissue that, like other connective tissues, consists of cells and extracellular matrix. • The feature that distinguishes bone from other connective tissues is the mineralization of its matrix, which produces an extremely hard tissue capable of providing support and protection. • The mineral is calcium phosphate in the form of hydroxyapatite crystals • Bone matrix contains mainly type I collagen along with other matrix (noncollagenous) proteins. Bone matrix contains lacunae connected by a network of canaliculi. Within the bone matrix are spaces called lacunae (sing., lacuna), each of which contains a bone cell, or osteocyte. The osteocyte extends numerous processes into small tunnels called canaliculi. In addition to osteocytes, four other cell types are associated with bone. • Osteoprogenitor cells are cells derived from mesenchymal stem cells; they give rise to osteoblasts. • Osteoblasts are cells that secrete the extracellular matrix of bone; once the cell is surrounded with its secreted matrix, it is referred to as an osteocyte. • Bone-lining cells are cells that remain on the bone surface when there is no active growth. They are derived from those osteoblasts that remain after bone deposition ceases. • Osteoclasts are bone-resorbing cells present on bone surfaces where bone is being removed or remodeled (reorganized) or where bone has been damaged. Bones are the organs of the skeletal system; bone tissue is the structural component of bones. • Typically, a bone consists of bone tissue and other connective tissues, including hemopoietic tissue, fat tissue, blood vessels, and nerves. • If the bone forms a freely movable or synovial joint, hyaline cartilage is present. The ability of the bone to perform its skeletal function is attributable to the bone tissue and, where present, the hyaline or articular cartilage. • Bone tissue is classified as either compact (dense) or spongy (cancellous). Bones are classified according to shape; the location of spongy and compact bone varies with bone shape. Spongy and compact bone tissues are located in specific parts of bones. It is useful, then, to outline briefly the kinds of bones and survey where the two kinds of bone tissue are located. On the basis of shape, bones can be classified into four groups: • Long bones are longer in one dimension than other bones and consist of a shaft and two ends (e.g., the tibia and the metacarpals). • Short bones are nearly equal in length and diameter (e.g., the carpal bones of the hand). • Flat bones are thin and platelike (e.g., the bones of the calvaria and the sternum). They consist of two layers of relatively thick compact bone with an intervening layer of spongy bone. • Irregular bones have a shape that does not fit into any one of the three groups just described; the shape may be complex (e.g., a vertebra), or the bone may contain air spaces or sinuses (e.g., the ethmoid bone). Long bones have a shaft, called the diaphysis, and two expanded ends, each called an epiphysis. The articular surface of the epiphysis is covered with hyaline cartilage. The flared portion of the bone between the diaphysis and the epiphysis is called the metaphysis. A large cavity filled with bone marrow, called the marrow or medullary cavity, forms the inner portion of the bone. Short bones possess a shell of compact bone and have spongy bone and a marrow space on the inside. Elsewhere, periosteum, a fibrous connective tissue capsule covers the outer surface of the bone. BONE FORMATION The development of a bone is traditionally classified as endochondral or intramembranous. The distinction between endochondral and intramembranous formation rests on whether a cartilage model serves as the precursor of the bone (endochondral ossification) or whether the bone is formed by a simpler method, without the intervention of a cartilage precursor intramembranous ossification). The bones of the extremities and those parts of the axial skeleton that bear weight (e.g., vertebrae) develop by endochondral ossification. The flat bones of the skull and face, the mandible, and the clavicle develop by intramembranous ossification. Schematic diagram of developing long bone. PHYSIOLOGIC ASPECTS OF BONE • Bone serves as a reservoir for body calcium. The maintenance of normal blood calcium levels is critical to health and life. Calcium may be delivered from the bone matrix to the blood if the circulating blood levels of calcium fall below a critical point (physiologic calcium concentration in the human ranges from 8.9 to 10.1 mg/dL). Conversely, excess blood calcium may be removed from the blood and stored in bone. These processes are regulated by parathyroid hormone (PTH), secreted by the parathyroid gland, and calcitonin, secreted by the parafollicular cells of the thyroid gland . PTH acts on the bone to raise low blood calcium levels to normal. Calcitonin acts to lower elevated blood calcium levels to normal. • Bone can repair itself after injury. The initial response to a fracture is similar to the response to any injury that produces tissue destruction and hemorrhage. New loose connective tissue, granulation tissue, is formed, and as this tissue becomes denser, cartilage forms in parts of it. Cartilage is a form of connective tissue composed of cells called chondrocytes and a highly specialized extracellular matrix. • Cartilage is an avascular tissue that consists of chondrocytes and an extensive extracellular matrix. More than 95% of cartilage volume consists of extracellular matrix, which is a functional element of this tissue. The chondrocytes are sparse but essential participants in producing and maintaining the matrix. • The extracellular matrix in cartilage is solid and firm but also somewhat pliable, which accounts for its resilience. Three types of cartilage that differ in appearance and mechanical properties are distinguished on the basis of characteristics of their matrix: Hyaline cartilage is characterized by matrix containing type II collagen fibers, GAGs, proteoglycans, and multiadhesive glycoproteins. Elastic cartilage is characterized by elastic fibers and elastic lamellae in addition to the matrix material of hyaline cartilage. Fibrocartilage is characterized by abundant type I collagen fibers as well as the matrix material of hyaline cartilage. HYALINE CARTILAGE • Hyaline cartilage is distinguished by a homogeneous, amorphous matrix. • The matrix of hyaline cartilage appears glassy in the living state: hence, the name hyaline [Gr. hyalos, glassy]. Throughout the cartilage matrix are spaces called lacunae. Located within these lacunae are the chondrocytes. • Hyaline cartilage is not a simple, inert, homogeneous substance but a complex living tissue. It provides a low-friction surface, participates in lubricating synovial joints, and distributes applied forces to the underlying bone. ELASTIC CARTILAGE Elastic cartilage is distinguished by the presence of elastin in the cartilage matrix. • In addition to containing the normal components of hyaline cartilage matrix, elastic cartilage matrix also contains a dense network of branching and anastomosing elastic fibers and interconnecting sheets of elastic material. • Elastic cartilage is found in the external ear, the walls of the external acoustic meatus, the auditory (Eustachian) tube, and the epiglottis of the larynx. The cartilage in all of these locations is surrounded by a perichondrium similar to that found around most hyaline cartilage. Unlike hyaline cartilage, which calcifies with aging, the matrix of elastic cartilage does not calcify during the aging process. FIBROCARTILAGE Fibrocartilage consists of chondrocytes and their matrix material in combination with dense connective tissue. • Fibrocartilage is a combination of dense regular connective tissue and hyaline cartilage. The chondrocytes are dispersed among the collagen fibers singularly, in rows, and in isogenous groups. • These chondrocytes appear similar to the chondrocytes of hyaline cartilage, but they have considerably less cartilage matrix material. • There is also no surrounding perichondrium as in hyaline and elastic cartilage. • Fibrocartilage is typically present in intervertebral discs, the symphysis pubis, articular discs of the sternoclavicular and temporomandibular joints, menisci of the knee joint, the triangular fibrocartilage complex of the wrist, and certain places where tendons attach to bones. CHONDROGENESIS AND CARTILAGE GROWTH Most cartilage arises from mesenchyme during chondrogenesis. Chondrogenesis, the process of cartilage development, begins with the aggregation of chondroprogenitor mesenchymal cells to form a mass of rounded, closely apposed cells. In the head, most of the cartilage arises from aggregates of ectomesenchyme derived from neural crest cells. The site of hyaline cartilage formation is recognized initially by an aggregate of mesenchymal or ectomesenchymal cells known as a chondrogenic nodule. Cartilage has limited ability for repair. When hyaline cartilage calcifies, it is replaced by bone. • The portion of articular cartilage that is in contact with bone tissue in growing and adult bones, but not the surface portion, is calcified. • Calcification always occurs in cartilage that is about to be replaced by bone (endochondral ossification) during an individual’s growth period. • Hyaline cartilage in the adult calcifies with time as part of the aging process. Cartilage is capable of two kinds of growth, appositional and interstitial. With the onset of matrix secretion, cartilage growth continues via a combination of two processes: • Appositional growth, the process that forms new cartilage at the surface of an existing cartilage; and • interstitial growth, the process that forms new cartilage within an existing cartilage mass Muscle Tissue Muscle tissue is responsible for movement of the body and its parts and for changes in the size and shape of internal organs. This tissue is characterized by aggregates of specialized, elongated cells arranged in parallel array that have the primary role of contraction. Myofilament interaction is responsible for muscle cell contraction. Two types of myofilaments are associated with cell contraction. •Thin filaments (6 to 8 nm in diameter, 1.0 m long) are composed primarily of the protein actin. Each thin filament of fibrous actin (Factin) is a polymer formed from globular actin molecules (G-actin). • Thick filaments (15 nm in diameter, 1.5 m long) are composed of the protein myosin II. Each thick filament consists of 200 to 300 myosin II molecules. The long, rodshaped tail portion of each molecule aggregates in a regular parallel but staggered array, whereas the head portions project out in a regular helical pattern. The two types of myofilaments occupy the bulk of the cytoplasm, which in muscle cells is also called sarcoplasm [Gr. sarcos, flesh; plasma, thing ]. Actin and myosin are also present in most other cell types (although in considerably smaller amounts), where they play a role in cellular activities such as cytokinesis, exocytosis, and cell migration. In contrast, muscle cells contain a large number of aligned contractile filaments that the cells use for the single purpose of producing mechanical work. Muscle is classified according to the appearance of the contractile cells. Two principal types of muscle are recognized: Striated muscle, in which the cells exhibit crossstriations at the light microscope level, and Smooth muscle, in which the cells do not exhibit crossstriations. Striated muscle tissue is further subclassified on the basis of its location: • Skeletal muscle is attached to bone and is responsible for movement of the axial and appendicular skeleton and for maintenance of body position and posture. In addition, skeletal muscles of the eye (extraocular muscles) provide precise eye movement. • Visceral striated muscle is morphologically identical to skeletal muscle but is restricted to the soft tissues, namely, the tongue, pharynx, lumbar part of the diaphragm, and upper part of the esophagus. These muscles play essential roles in speech, breathing, and swallowing. • Cardiac muscle is a type of striated muscle found in the wall of the heart and in the base of the large veins that empty into the heart. SKELETAL MUSCLE • A skeletal muscle cell is a multinucleated syncytium. In skeletal muscle, each muscle cell, more commonly called a muscle fiber, is actually a multinucleated syncytium. A muscle fiber is formed during development by the fusion of small, individual muscle cells called myoblasts. • The nuclei of a skeletal muscle fiber are located in the cytoplasm immediately beneath the plasma membrane, also called the sarcolemma. • A skeletal muscle consists of striated muscle fibers held together by connective tissue. • The connective tissue that surrounds both individual muscle fibers and bundles of muscle fibers is essential for force transduction. The connective tissue associated with muscle is named according to its relationship with the muscle fibers: • Endomysium is the delicate layer of reticular fibers that immediately surrounds individual muscle fibers. Only smalldiameter blood vessels and the finest neuronal branches are present within the endomysium, running parallel to the muscle fibers. • Perimysium is a thicker connective tissue layer that surrounds a group of fibers to form a bundle or fascicle. Fascicles are functional units of muscle fibers that tend to work together to perform a specific function. Larger blood vessels and nerves travel in the perimysium. • Epimysium is the sheath of dense connective tissue that surrounds a collection of fascicles that constitutes the muscle. The major vascular and nerve supply of the muscle penetrates the epimysium. • Three types of skeletal muscle fibers - red, white, and intermediate - can be identified by color in vivo. • Skeletal muscle fibers are characterized by speed of contraction, enzymatic velocity, and metabolic activity. • The three types of skeletal muscle fibers are type I (slow oxidative), type IIa (fast oxidative glycolytic), and type Iib (fast glycolytic) fibers. Three types of fiber are typically found in any given skeletal muscle; the proportion of each type varies according to the functional role of the muscle. Myofibrils and Myofilaments • The structural and functional subunit of the muscle fiber is the myofibril (A muscle fiber is filled with longitudinally arrayed structural subunits called myofibrils). • Myofibrils are composed of bundles of myofilaments. Myofilaments are the individual filamentous polymers of myosin II (thick filaments) and actin and its associated proteins (thin filaments). Myofilaments are the actual contractile elements of striated muscle. The bundles of myofilaments that make up the myofibril are surrounded by a well- developed smooth-surfaced endoplasmic reticulum (sER), also called the sarcoplasmic reticulum. Cross-striations are the principal histologic feature of striated muscle. • Cross-striations are evident in H&E–stained preparations of longitudinal sections of muscle fibers. They may also be seen in unstained preparations of living muscle fibers examined with a phase contrast or polarizing microscope, in which they appear as alternating light and dark bands. These bands are termed the A band and the I band. In polarizing microscopy, the dark bands are birefringent (i.e., they alter the polarized light in two planes). • Therefore, the dark bands, being doubly refractive, are anisotropic and are given the name A band. The light bands are monorefringent (i.e., they do not alter the plane of polarized light). Therefore, they are isotropic and are given the name I band. • Both the A and I bands are bisected by narrow regions of contrasting density. • The light I band is bisected by a dense line, the Z line, also called the Z disc. • The dark A band is bisected by a less dense, or light, region called the H band. • Furthermore, bisecting the light H band is a narrow dense line called the M line. The functional unit of the myofibril is the sarcomere, the segment of the myofibril between two adjacent Z lines. • The sarcomere is the basic contractile unit of striated muscle. It is the portion of a myofibril between two adjacent Z lines. • A sarcomere measures 2 to 3 µm in relaxed mammalian muscle. It may be stretched to more than 4 µm and, during extreme contraction, may be reduced to as little as 1 µm . The entire muscle cell exhibits crossstriations because sarcomeres in adjacent myofibrils are in register. The events leading to contraction of skeletal muscle can be summarized as a series of steps. The events involved in contraction can be summarized as follows: 1. The contraction of a skeletal muscle fiber is initiated when a nerve impulse traveling along the axon of a motor neuron arrives at the neuromuscular junction. 2. The nerve impulse prompts the release of acetylcholine into the synaptic cleft that binds into ACh-gated Na+ channels causing local depolarization of sarcolemma. 3. Voltage-gated Na + channels open, and Na + enters the cell. 4. General depolarization spreads over the plasma membrane of the muscle cell and continues via membranes of the T tubules. 5. Voltage sensor proteins in the plasma membrane of T tubules change their conformation. 6. At the muscle cell triads, the T tubules are in close contact with the lateral enlargements of the sarcoplasmic reticulum, where gated Ca2 + -release channels are activated by conformational changes of voltage-sensor proteins. 7. Ca2 + is rapidly released from the sarcoplasmic reticulum into the sarcoplasm. 8. Ca2 + binds to the TnC portion of the troponin complex. 9. The contraction cycle is initiated, and Ca2 + is returned to the terminal cisternae of the sarcoplasmic reticulum. CARDIAC MUSCLE Cardiac muscle has the same types and arrangement of contractile filaments as skeletal muscle. Therefore, cardiac muscle cells and the fibers they form exhibit crossstriations evident in routine histologic sections. In addition, cardiac muscle fibers exhibit densely staining crossbands, called intercalated discs, that cross the fibers in a linear fashion or frequently in a way that resembles the risers of a stairway Structure of Cardiac Muscle • The cardiac muscle nucleus lies in the center of the cell. • Numerous large mitochondria and glycogen stores are adjacent to each myofibril. • The intercalated discs represent junctions between cardiac muscle cells. • The sER in cardiac muscle cells is organized into a single network along the sarcomere, extending from Z line to Z line. • Passage of Ca2+ from the lumen of the T tubule to the sarcoplasm of a cardiac muscle cell is essential to initiate the contraction cycle. • Cardiac muscle cells exhibit a spontaneous rhythmic contraction. SMOOTH MUSCLE • Smooth muscle generally occurs as bundles or sheets of elongated fusiform cells with finely tapered ends. • The cells, also called fibers, range in length from 20 µm in the walls of small blood vessels to about 200 µm in the wall of the intestine; they may be as large as 500µ m in the wall of the uterus during pregnancy. Smooth muscle cells are interconnected by gap junctions, the specialized communication junctions between the cells. Structure of Smooth Muscle • Smooth muscle cells possess a contractile apparatus of thin and thick filaments and a cytoskeleton of desmin and vimentin intermediate filaments. • Dense bodies provide an attachment site for thin filaments and intermediate filaments. • Contraction in smooth muscles is initiated by a variety of impulses, including mechanical, electrical, and chemical stimuli. • Smooth muscle cells lack a T system. • Contraction of smooth muscle is regulated by the Ca2+– calmodulin–myosin light chain kinase system. • The force of smooth muscle contraction may be maintained for long periods in a “latch state.” • Smooth muscle is specialized for slow, prolonged contraction.