Alternation of generations: a review
advertisement

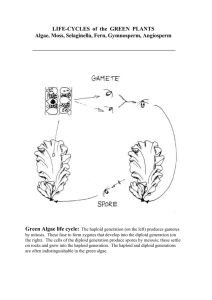
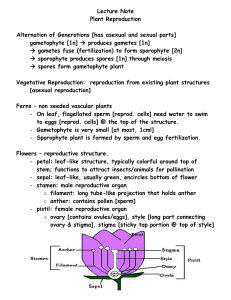
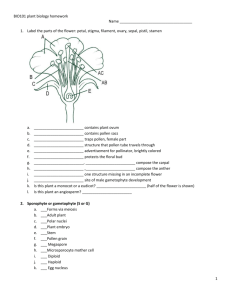
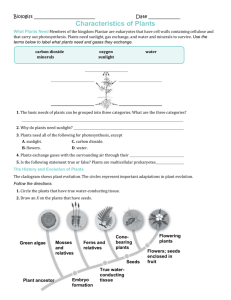
Plant Reproduction Level 1 Biological Diversity Jim Provan Campbell: Chapter 38 Alternation of generations: a review Angiosperm life cycle includes alternation of generations: haploid gametophyte generations alternate with diploid sporophyte generation: Sporophyte is recognisable “plant” - produces haploid spores by meiosis in sporangia Spores undergo mitotic division and develop into multicellular male or female gametophyte Gametophytes produce gametes (sperm and eggs) by mitosis: gametes fuse to form zygote which develops into multicellular sporophyte Sporophyte is dominant in angiosperm life cycle: gametophyte stage is reduced and is totally dependent on sporophyte Alternation of generations: a review Variations on the basic flower structure Complete: has sepals, petals, stamens and carpels Incomplete: missing one or more organs (e.g. grasses) Perfect: has both stamens and carpels Imperfect: either staminate or carpellate - unisex Monoecious: has both staminate and carpellate flowers on same plant Dioecious: has staminate and carpellate flowers on separate individual plants Floral diversity Development of male gametophyte (pollen) Within sporangial chamber of anther, diploid microsporocytes undergo meiosis to form four haploid microspores Haploid microspore nucleus undergoes mitotic division to give rise to a generative cell and a tube cell Wall of microspore thickens Development of female gametophyte (embryo sac) Megasporocyte in sporangium of each ovule grows and goes through meiosis to form four haploid megaspores (only one usually survives) Remaining megaspore grows and its nucleus undergoes three mitotic divisions, forming one large cell with eight haploid nucleii Membranes partition this into a multicellular embryo sac Pollination brings male and female gametophytes together Pollination: the placement of pollen onto the stigma of a carpel: Some plants use wind to disperse pollen Others interact with animals that transfer pollen directly Some plants self pollinate, but most cross-pollinate Most monoecious angiosperms have mechanisms to prevent selfing - maximises genetic variation: Stamens and carpels may mature at different times Structural arrangement of flower reduces chance that pollinator will transfer pollen between anthers and stigma of same plant Some plants are self-incompatible Genetic basis of self-incompatibility Based on S genes Many alleles in plant population gene pool Pollen landing on stigma with same allele at S-locus is self-incompatible: Pollen grain will not initiate or complete formation of pollen tube Prevents fertilisation between plants with similar S-alleles Multiple mechanisms at S-loci Mechanism underlying inhibition of pollen tube varies: Block occurs in pollen grain (gametophyte selfincompatibility): RNAses from carpel enter pollen and destroy RNA Block occurs in stigma (sporophyte selfincompatibility) e.g. signal transduction systems in mustards Double fertilisation gives rise to the zygote and the endosperm Double fertilisation: union of two sperm cells with two cells of the embryo sac Pollen grain germinates and extends pollen tube Generative cell undergoes mitosis, forming two sperm Pollen tube enters through micropyle and discharges sperm One sperm unites with egg Other sperm unites with polar nuclei forming endosperm (3n) Endosperm development Begins before embryo development Triploid nucleus divides to form multinucleate “supercell” This undergoes cytokinesis, forming cell membranes and walls and thus becoming multicellular: Endosperm is rich in nutrients, which it provides to the developing embryo In most monocots, endosperm stocks nutrients that can be used by the seedling after germination In many dicots, food reserves of the endosperm are exported to the cotyledons Embryo development First mitotic division transverse: Large basal cell forms suspensor Terminal cell divides several times to form spherical proembryo Cotyledons appear at either side of apical meristem Suspensor attaches at apex of embryonic root and meristem After germination, apical and root meristems sustain growth Structure of the mature seed In dicot seeds: Hypocotyl terminates in the radicle (embryonic root) Epicotyl terminates in the plumule (shoot tip) Monocot seeds have a special cotyledon called a scutellum: Large surface area - absorbs nutrients from endosperm during germination Embryo enclosed in sheath: - Coleoptile protects the shoot - Coleorhiza protects the root The ovary develops into a fruit adapted for seed dispersal A true fruit is a ripened ovary Fruits can be classified by their origin: Simple fruits: derived from a single ovary e.g. cherry Aggregate fruits: derived from a single flower with several carpels e.g. blackberry Multiple fruits: develop from an inflorescence Seed dormancy Prevents germination when conditions for seedling growth are unfavourable Conditions for breaking dormancy vary depending on type of environment plant occupies: Seeds of desert plants will not germinate until there has been a heavy rainfall and not after a light shower In chaparral regions where bushfires are common, seeds may not germinate until exposed to heat of fire which clears away older, competing vegetation Other seeds require cold, sunlight or passage through an animal’s digestive system Viability ranges from a few days to decades Seed germination Imbibition causes seed to swell, rupturing seed coat Metabolic changes restart growth of the embryo Storage materials are digested by enzymes and nutrients transferred to growing parts of embryo Radicle (embryonic root) emerges from seed Seed germination (continued) Next stage involves shoot tip breaking through soil surface: In many dicots, a hook forms in the hypocotyl Light stimulates the hypocotyl to straighten, raising the cotyledons Other plant species follow different germination methods: In peas, hook forms in epicotyl which straightens and leaves cotyledons in ground In monocots, shoot grows straight up through coleoptile tube Many plants can clone themselves by asexual reproduction Asexual reproduction: production of offspring from a single parent without recombination clones Two natural mechanisms of vegetative reproduction: Fragmentation: separation of parent plant into parts that reform whole plants: - Most common form of vegetative reproduction - Some species of dicots develop adventitious shoots that become separate shoot systems Apomixis: production of seeds without meiosis and fertilisation: - Diploid cell in ovule gives rise to an embryo - Ovules mature into seeds which are dispersed (e.g. dandelions) Sexual and asexual reproduction are complementary in many plants Both have had featured roles in adaptation of plant populations to their environments Benefits of sexual reproduction: Generates genetic variation Produces seeds, which can disperse to new locations Benefits of asexual reproduction: In a stable environment, plants can clone many copies of themselves in a short period Progeny are mature fragments of the parental plant, as opposed to small, fragile seedlings produced by sexual reproduction