22-Cerebellar Anatomy

NBIO 401 Fall 2013
Cerebellar Anatomy
Class 22– Friday, November 15, 2013
Robinson
Objectives :
- Be able to describe the position of the cerebellum in the brain physically and how it connects to other parts of the brain
- Be able to characterize the major parts of the cerebellum and what is distinctive about them
- Be able to describe the cerebellar nuclei and their connections to the cerebellar cortex
- Be able to explain the structure of the cerebellar cortex and the two pathways through it
- Be able to characterize the relationship between the inferior olive and the cerebellum
Cerebellum and Its Parts
The cerebellum is the large folded structure apparently wedged between the top of the brainstem and the posterior part of the cerebral cortex. The bottom of the cerebellum forms the 4th ventricle. Though apparently smaller than the cerebral cortex, the cerebellum contains ~2X as many neurons as the cerebral cortex. Its function is to make our movements as fast, accurate, smooth, and consistent as possible. Cerebellar damage does not cause paralysis but renders movements slow, inaccurate, inconsistent, and jerky. Though the cerebellum sends very few fibers to the spinal cord it exerts a powerful influence on movements via projections, either direct or via one relay neuron, to the structures from which all four major descending motor tracks originate.
As the picture below shows, the smallest folds, which form the ridges visible on the outside of the cerebellum, are called folia. Groups of adjacent folia form lobules that are divided by fissures.
-Class 22, page 1-
NBIO 401 Fall 2013
We number the with roman numerals lobules I-X. As the picture below shows, the lobule just rostral of the 4 th
ventricle is I and the one just caudal to it is X. Between lobules V and VI the primary fissure separates the posterior lobe from the anterior lobe.
As the picture below shows, the anterior lobe receives its input primarily from the spinal cord and is referred to as the paleocerebellum.
The posterior lobe receives its input primarily from the cerebral cortex via relay neurons in the pontine nuclei and is called the neocerebellum.
Between lobules IX and X, the posteriorlateral fissure separates the posterior lobe from the flocculonodular lobe. This lobe receives most of its input from the vestibular system and is called the vestibulocerebellum.
-Class 22, page 2-
NBIO 401 Fall 2013
In this view looking caudally and ventrally at the cerebellum after removing the cerebral cortex, we see the position of the primary fissure with the anterior lobe rostrally and the posterior lobe caudally. The region near the midline that is outlined in pink is the vermis. This name is from the Latin for worm because it resembles a segmented worm.
The lateral extensions of the cerebellum are called the hemispheres. Note how large the hemispheres and the posterior lobe are.
These structures receive input from and send signals to the cerebral cortex. In animals like humans with a large cerebral cortex the posterior lobe and hemispheres are very large.
To summarize locations in the cerebellum it is common to use drawings like the one below on the left. Note that the lobules are labeled along the vermis.
The human cerebellum if unrolled would look approximately like this>
The numbered lobules extend laterally into the hemispheres and are called the hemispheric parts of the lobules e.g., hemispheric VI or HVI. As labeled on the right side of the drawing. The left side of the drawing shows the name of the lateral parts of the cerebellum in an older naming system, sometimes still used by clinicians.
The drawing on the left is useful but distorts the true shape of the cerebellar cortex.
The drawing on the right shows the approximate shape of a human cerebellar cortex it were unrolled.
-Class 22, page 3-
NBIO 401
LEG
Somatotopy in the cerebellum is not as clearly organized and orderly as that in motor and somatosensory cortex . Nonetheless, it is clear that particular parts of the cerebellum influence the movements of particular parts of the body. Right is a view of the cerebellum from above and behind that shows regions associated with particular parts of the body. The region marked as receiving visual and auditory signals influences eye movements.
VISUAL &
AUDITORY
HEAD
ARM
HEAD
ARM
LEG
Cerebellum Connections to the Rest of the Brain
The multiple inputs to the cerebellum combine in a way that we do not understand yet. The resultant signals are carried in the activity of the neurons in the cerebellar nuclei.
The axons of these neurons form the superior cerebellar peduncle (see below) and terminate mainly in the four structures that send tracts down the spinal cord to influence spinal motoneurons.
These structures include:
1) the motor regions of the cerebral cortex (contacted via a relay in the thalamus)
2) the red nucleus
3) the vestibular nuclei
4) the reticular nuclei
Three large fiber bundles on each side of the cerebellum connect the cerebellum to the rest of the brain. These large bundles are called peduncles. A is a schematic representation of the cerebellum as viewed above and behind showing the general position of the three peduncles on each
-Class 22, page 4-
Fall 2013
NBIO 401 Fall 2013 side. The inferior and middle peduncles carry fibers into the cerebellum. The superior peduncle carries fibers leaving the cerebellum. There are ~40X as many fibers coming into the cerebellum as leaving it.
The inferior peduncle (restiform body in older terminology) carries fibers originating in the spinal cord, vestibular labyrinth, and vestibular nuclei into the cerebellum. The middle peduncle (brachium pontis) consists of the axons of neurons in the pontine nuclei relaying information from the cerebral cortex to the cerebellum. The superior peduncle (brachium conjunctivum) carries fibers coming out of the cerebellum. Fibers in the superior peduncle terminate in motor-related brain structures. B shows the positions of the peduncles in a human brain. C shows the cut surfaces of the peduncles in a human brain after we have removed the cerebellum from the brainstem.
Below we will examine the two peduncles that bring most input into the cerebellum, the inferior and middle cerebellar peduncles. Next we will describe the paths that incoming information takes through the cerebellum. Finally we will look at the targets of cerebellar output.
Inferior
Cerebellar
Peduncle
These two transverse sections through the medulla show the inputs reaching the cerebellum through the inferior cerebellar peduncle. The inferior cerebellar peduncle is a large fiber bundle in the extreme dorsolateral part of the medulla. The constituents of the inferior cerebellar peduncles include both climbing fibers and mossy fibers. The climbing fibers originate in the contralateral inferior olive. The large folded part of the inferior olive shown here is the principle nucleus. The two much smaller nuclei of the inferior olive, the medial and dorsal accessory nuclei, are visible medial to the principle nucleus. They carry somatosensory and proprioceptive signals, respectively, from the contralateral body and head. Mossy fibers from the spinal cord and external cuneate nucleus carry proprioceptive information from the legs and arms, respectively, to the ipsilateral cerebellum. Mossy fibers from the vestibular and reticular nuclei carry information about head and body movement in space to the ipsilateral cerebellum.
-Class 22, page 5-
NBIO 401 Fall 2013
Inferior Peduncle – Mossy Fibers
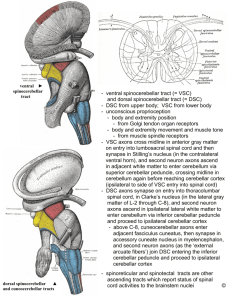
The cerebellum receives proprioceptive information from the spinal cord via two tracts, the dorsal spinocerebellar tract (DSCT) and the ventral spinocerebellar tract (VSCT).
The information in both pathways is unconscious, i.e., a lesion severing these tracts results in no loss of sensation. Such lesions make movements of the affected limb clumsy.
The DSCT originates in the nucleus dorsalis of Clarke, or Clarke’s column. This column of cells exists
ONLY in the THORACIC and UPPER
LUMBAR levels of the spinal cord ( T1
- L2 ). It consists of a column of large round neurons in the ventral-medial region of the dorsal horn.
Input to Clarke’s column originates with neurons in the ipsilateral dorsal root ganglion (DRG) that carry proprioceptive, touch, and pressure information. Axons originating in DRGs caudal to the thoracic cord, i.e., in lumbar or sacral cord segments (carrying leg information) course up the dorsal columns to the thoracic cord, where they terminate. Axons originating in thoracic DRGs terminate on Clarke’s column cells in their segment. Axons originating in Clarke’s column travel laterally to the extreme dorsolateral edge of the cord and ascend directly into the cerebellum via the inferior cerebellar peduncle.
Axons originating in DRGs above thoracic segments (carrying arm information) travel up the dorsal columns to the external cuneate nucleus (ECN) and terminate. Thus the ECN is analogous to Clarke’s column for the arm. Axons originating from cells in the ECN travel laterally, join the fibers from Clarke’s column, and enter the cerebellum via the inferior peduncle. This pathway is called the cuneocerebellar tract. Note that no component of the
DSCT ever crosses.
The VSCT originates in cells outside Clarke’s column. Their axons cross and ascend contralaterally to the superior cerebellar peduncle where they enter and cross again to terminate in the cerebellum on the side from which they originated. We know less about information carried by the VSCT. Like the DSCT, its fibers carry proprioception, but it seems to be more related to movements of entire limbs, not just of one joint or muscle like DSCT fibers.
-Class 22, page 6-
NBIO 401
Terminations of mossy fibers, like those coming from the spinal cord, in the anterior lobe of a cat cerebellar cortex.
Note that they can occupy very well defined regions of the cortex.
Inferior Peduncle – Climbing Fibers
Fall 2013
Climbing fibers are fibers from the inferior olive that enter the cerebellum through the inferior cerebellar peduncle and terminate in the cerebellar cortex. They terminate in parasagittal stripes that are clearly visible in A , a frontal section through the anterior lobe of a cat cerebellum. The axons whose terminals form each stripe originate in a different part of the inferior olive (IO). B is an unrolled map of the cerebellar cortex of a cat showing the position of different IO terminal stripes in the top part of the diagram. In the bottom part is a map of the three divisions of the IO showing the location of neurons whose axons form each stripe. The thick horizontal stripe in the upper half of the drawing shows the approximate level of the section in A . The two stripes visible in A are the two stripes, C1 and C3, which originate in the dorsal accessory olive.
-Class 22, page 7-
NBIO 401 Fall 2013
The top transverse section shows the large cerebral peduncle carrying fibers descending from the cerebral cortex. The overwhelming majority of these fibers terminate in the pontine nuclei, not the spinal cord. The bottom two sections are through the pontine nuclei. The superimposed drawings show that fibers from the cerebral cortex terminate in the ipsilateral pontine nuclei.
The pontine neurons receiving this input send their axons across the midline into the middle cerebellar peduncle. The fibers terminate as mossy fibers in the granular layer of the contralateral cerebellar cortex.
Inside the Cerebellum
In a transverse section through the cerebellum we can see that it consists of two components, the cerebellar cortex visible on the outside and the cerebellar nuclei embedded within the white matter under the cortex. The white matter consists of fibers coming into the cerebellum and the axons of the output neurons of the cerebellar cortex, Purkinje cells, coursing to terminate in the cerebellar nuclei.
There are four nuclei on each side. Most laterally is the dentate or lateral nucleus. Most medially is the fastigial or medial nucleus. Between the lateral and medial nuclei are the intermediate, or interpositus nuclei, the globose (more lateral) and emboliform (more medial).
Generally each cerebellar nucleus receives input via Purkinje cell axons from the cortex that is nearest to it. Thus the lateral nucleus receives input from the lateral cortex in the cerebellar hemispheres, the medial nucleus from the medial cortex of the vermis, and the intermediate nuclei from intermediately placed cortex. The cerebellar nuclei are the output structures of the cerebellum. The axons of neurons in the cerebellar nuclei form the superior cerebellar peduncle.
-Class 22, page 8-
NBIO 401 Fall 2013
Looking into a transparent cerebellum in posterior view we see the positions of the cerebellar nuclei relative to each other and the cerebellar cortex. Note how much larger the dentate nucleus is than the other nuclei. Higher animals that have a large cerebral cortex also have a large dentate nucleus, large lateral cerebellar cortex, and large pontine nuclei. This makes sense because the cerebral cortex projects to the pontine nuclei which project, in turn, mostly to the lateral cerebellar cortex, whose output then reaches the dentate nucleus. There is another structure, associated with these four, that is also large in higher animals: the principle nucleus of the inferior olive. This nucleus supplies a particular kind of input, climbing fibers, to the lateral cerebellar cortex.
Structure of Cerebellar Cortex
Here we see the three layers of the cerebellar cortex and the connections between neurons in these layers. Incoming mossy fibers terminate on the dendrites of granular cells. Granular cell axons course directly toward the surface and bifurcate, sending one branch in each direction for ~ 2-6 mm. These branches are parallel to those of other granular cells and to the long axis of the folium and are called parallel fibers.
As the insert in the upper left shows, this means that most parallel fibers run perpendicular to the long axis of the body. Parallel fibers contact the dendrites of one Purkinje cell after another along their course. Purkinje cell dendrites are broad and flat, like a hand with outstretched fingers. The broad dimension is oriented perpendicular to the parallel fibers so that a Purkinje cell receives input from ~100,000 – 300,000 of them. Purkinje cell axons run through the white matter to terminate in the cerebellar nuclei. All output from the cerebellar cortex leaves the cortex via Purkinje cell axons. Climbing fibers terminate directly on the dendrites of ~10
Purkinje cells. An action potential in a climbing fiber always causes an action potential in the
Purkinje cells that it contacts. Thus a mossy fiber has a small effect on many Purkinje cells, whereas climbing fibers have a large effect on a small number of Purkinje cells. Finally, there are three kinds of interneurons in the cerebellar cortex; stellate and basket cells in the inner and outer parts of the molecular layer, respectively, and Golgi cells in the granular cell layer.
-Class 22, page 9-
NBIO 401
Here we see two views of a cerebellar Purkinje cell. In A we see the cell as if we were looking along the long axis of the folium. The parallel fibers are coming out of the page directly at you. In B we see the cell as if we were looking at it from an angle perpendicular to the long axis of the folium. If you could see the parallel fibers in B , they would be running horizontally across the picture above the Purkinje cell soma. Note that the
Purkinje cell’s dendritic tree is much broader in A than in B .
To the right is a Nissl stained section showing the four layers of cerebellar cortex. Moving downward from the surface of the cortex, the molecular layer is first. It contains mostly fibers, Purkinje cell dendrites, and granular cell axons, with a few scattered cells. Under that is the
Purkinje cell layer, a single layer of
Purkinje cell bodies. Their dendrites extend upward into the molecular layer. Under Purkinje cells is the granular layer, a layer of small, densely packed cells. Mossy fibers terminate in the granular layer. It is granular cells that account for the astonishingly large number of neurons within the cerebellum. Beneath the granular layer is the white matter, consisting of incoming climbing and mossy fibers and Purkinje cell dendrites.
In histological sections of the cerebellar cortex we often see the layers arranged as in the slightly lower power picture to the right. This section is in a parasagittal plane (How do we know that by looking at this section?). We see the molecular layer on top, the densely stained granular layer beneath it and the
Purkinje cell layer between them. Under the granular layer we can see some white matter.
Fall 2013
-Class 22, page 10-
NBIO 401 Fall 2013
Above is a simplified view of the cerebellar cortex and the pathway through it taken by signals arriving on mossy fibers and climbing fibers.
Mossy Fibers - Incoming mossy fibers terminate on the dendrites of granule cells.
Granular cell axons form the parallel fibers in the molecular layer. Parallel fibers contact the dendrites of one Purkinje cell after another along their course. Purkinje cells receive input from ~100,000 parallel fibers. Thus an incoming signal integrates with many other signals in a
Purkinje cell. The resultant signal travels in the Purkinje cell axon through the white matter to the cerebellar nuclei. All output from the cerebellar cortex leaves the cortex via Purkinje cell axons and INHIBITS the cells on which it terminates.
Climbing Fibers Each climbing fiber terminates on the dendrites of ~10 Purkinje cells. Each Purkinje cell receives input from only one climbing fiber. An action potential in a climbing fiber always causes an action potential, called a complex spike, in the Purkinje cells that it contacts. Complex spikes occur at only 1 - 4 spikes/second and are accompanied by a very large influx of calcium ions into the Purkinje cell. The function of complex spikes is the subject of much interest. We will talk about it in more detail in a later lecture. Signals in complex spikes travel down Purkinje cell axons to neurons in the cerebellar nuclei. The significance of these signals is not yet clear.
Mossy fiber and climbing fiber input to a Purkinje cell elicit different types of action potentials in the Purkinje cell.
-Class 22, page 11-
NBIO 401 Fall 2013
Cerebellar Outpu t
After information from mossy fibers and climbing fibers converges in Purkinje cells, the axons of Purkinje cells convey the resulting signal, whose function we do not completely understand yet, to the cerebellar nuclei. The output of many Purkinje cells converges on a single neuron in the cerebellar nuclei.
The axons of cerebellar nuclear cells carry the output of the cerebellum, i.e., the signals that the cerebellum produces, to the motor structures from which the four major motor tracts originate, the motor cortex (via a relay in the thalamus), the red nucleus, the vestibular nuclei (not shown), and the reticular nuclei, (not shown).
Here is another representation of the pathway from the cerebellar nuclei to the motor structures from which the four major motor tracts originate, the motor cortex (via a relay in the
-Class 22, page 12-
NBIO 401 Fall 2013 thalamus), the red nucleus, the vestibular nuclei (not shown), and the reticular nuclei (not shown). A shows the pathway from the right dentate nucleus, via the thalamus and motor areas of the cerebral cortex, to the right side of the spinal cord. B shows the pathway from the interpositus nuclei to the right side of the spinal cord via the thalamus and motor cortex and also via the red nucleus.
-Class 22, page 13-
NBIO 401 Fall 2013
Below is a sagittal section of a cat brainstem and cerebellum. In white we see the pathway from the cerebellum to the red nucleus and then to the spinal cord. To show this pathway we injected the tracer WGA-HRP into the red nucleus on one side of the brain. This tracer travels in both the orthograde and retrograde directions in neurons. Neuron cell bodies in the anterior and posterior interpositus nuclei are labeled via retrograde transport from the red nucleus. The labeled axons of red nucleus neurons cross into the plane of the section and course through the brain stem to the spinal cord.
-Class 22, page 14-