Nolte – Chapter 20 – Cerebellum
advertisement

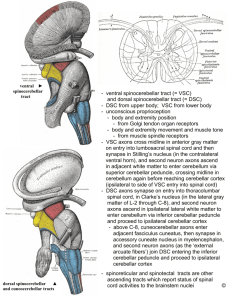
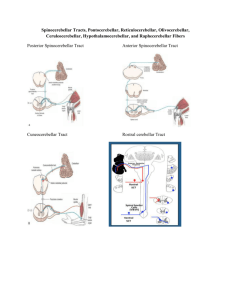
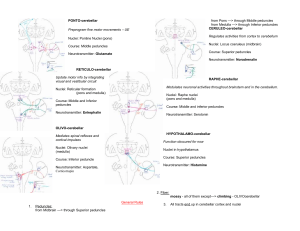
Nolte – Chapter 20 – Cerebellum There are cortical ridges all over the cerebellar surface known as folia The floculus of each side is (almost) pinched off in the front. The nodulus is the vermal portion of the floculonodular lobe and continue laterally into the floculus. The tonsils are the hemisphereal portions across the posterolateral fissure from the floccule. Their vermal continuation is the uvula. The primary fissure is a fissure that subdivides the body of the cerebellum into anterior and posterior lobes. The most medial zone, straddling the midline, is the vermis. On either side of the vermis is a cerebellar hemisphere with medial and lateral components. Inferior cerebellar peduncle brings in afferents from the spinal cord and brainstem (made up of the restiform and juxtarestiform(vestibular)) Middle cerebellar peduncle is the largest and emerges from the basal pons. Consists of afferents from the pontine nuclei on the contralateral side. The superior cerebellar peduncle consists of efferents from the cerebellum to the red nucleus and thalamus. Except the anterior(the one that relates to attempted movement and cross at the elvel of the spinal cord then goes to rostral pons, then comes back down)/rostral spinocerebellar tracts. The deep nuclei provide cerebellar output. The dentate nucleus is a crumpled sheet of cells that looks like the inferior olivary nucleus and most of the fibers in the superior cerebellar peduncle originate from the dentate gyrus. Gets its inputs mainly from the lateral cerebellar hemispheres. The interposed nucleus gets inputs from the medial hemisphere. The fastigial nucleus gets input from the vermis and then projects to the vestibular nuclei. The most superficial layer of the cerebellar cortex is the molecular layer Deep to the molecular later is a single layer of purkinje cells that sit with their axons sprawled in a “Mohawk” type fashion in a saggital view. Purkinjes are the only neurons whose axons leave the cerebellar cortex. adjacent to the medullary center is the granular layer. Each granule cell send its axon to the molecular layer where it bifurcates to form a fine, unmyleinated parallel fiber that extends along the long axis of the folium and synapses on many purkinjes. The afferent fibers to the cerebellar cortex are the climbing fibers and the mossy fibers. A single climbing fiber ends directly on one purkinje cell like ivy, climbing up the purkinje’s dendrites. It makes tens of thousands of excitatory synapses, making it the most powerful excitatory input in the entire brain. All of these climbing fibers arise in the contralateral inferior olive of the medulla and enter through the inferior cerebellar peduncle. The rest of the afferents are the mossy fibers which end on the dendrites of granule cells in an indirect route to the purkinje cells. Purkinje cells axons project to the deep nuclei and some actually leave without hitting these nuclei. The ones that don’t go to the deep nuclei exit via the juxtarestiform and end in the vestibular nuclei. Climbing fibers and mossy fibers also send collateral branches to the deep nuclei. These latter inputs seem to have a tonic excitatory drive for the nuclei and purkinje modulate this firing rate through inhibition. The flocculonodulat lobe and uvula receive vestibular inputs the vermis and medial hemispheres receive spinal inputs (somatosensory) from the spinocerebellar tracts and the cuneocerebellar tracts (caudal and rostral respectively). The trigeminal inputs from the head also come into the cerebellum. The anterior spinocerebellar(the one concerned with attempted movements that synapses immediately and then goes to rostral pons and comes back) enters through the superior cerebellar peduncle. All the rest of the somato cerebellar input goes through the inferior cerebellar peduncle. the lateral hemispheres get projections from cortex. Juxtarestiform inputs (vestibular) end as mossy fibers in the nodulus and uvula. The cerebral cortex projects to the cerebellum by way of the pontine nuclei, which get their inputs from the cerebral peduncle, which then cross the midline and enter the middle cerebellar peduncle. This is known as the corticocerebellar pathway. The vermis and medial hemispheres will get input from motor cortex and lateral portions will get premotor and somatosensory. Visual information from the superior colliculus andreticular formation come in this way as well. most of the outputs fromt hecerebellum go out the superior cerebellar peduncle and climb rostrally to decussate in the midbrain and synapse on the red nuclei and head to the thalamus (VL/VA). The red nucleus has magnocellular(usually axons from the interposed nucleus go here) cells that give rise to the rubrospinal tract and parvocellular(usually the dentate nuclei axons go here) cells that project to the inferior olive. The lateral hemispheres are involved in planning movements. The cerebral cortex and cerebellum talk back and forth via these connections. The planning of movements can be modulated by cerebellum by acting on the motor cortex. The dentate gyrus will change its firing rate before voluntary movements occur. Damage here causes limb ataxia and coordination of voluntary movements is estranged along with changes in muscle tone. Patients with damage see “intention tremors” as they voluntarily reach for things. These problems aren’t seen in rhythmic movements. This region might also be important for motor movements necessary for speech. The medial hemisphere is involved in adjusting limb movements. Medial hemispheres have somatopically organized inputs from the motor cortex and spinal cord and go out via interposed to motor cortex and red nucleus. This allows it to influence spinal cord motor neurons through the corticospinal tract and the rubrospinal tract. Seems as though it compares commands. This makes sense since the anterior spinocerebellar comes here. It can issue correcting signals by way of the interposed nucleus, which fires during movement. Damage here causes postural instability and ataxia. The vermis includes the representation of the trunk conveyed by the spinocerebellar tracts and has outputs to the vestibular nueclie by way of the fastigal nuclei, so it seems to be concerned with postural adjustments and creating stereotyped movements for rhythmic modulation of the basic pattern of walking. The flocculus and vermis are involved in eye movements, especially slow eye movements and the maintenance of equilibrium. Damage here results in loss of equilibrium and difficulty with pursuit eye movements or with making accurate voluntary movements. The cerebellum as a whole is involved in motor learning and gain setting. For instance, with the VOR, if there is normally a gain of 1 (the eyes move as much as the head), but this response becomes inappropriate, then the cerebellum can adjust this gain. REmving the flocculus or part of the inferior olivary nucleu prevents these adaptive changes. Conditions responses can also take place in the cerebellum. Removing the interposed nucleus abolishes conditioned responsed to the ipsilateral eye after one has been learned. Lesioning the inferior olive prevents acquisition of conditioned responses. Also, olive lesions after learning allow for a fading of the learning, suggesting its role in establishment and sustainability of conditioned responses. The cerebellum is also involved in cognitive functions. The lateral hemisperheres get projections from association cortex via the corticopontine fibers. The dentate can project to the profrontal cortex by way of the dorsomedial nucleus of the thalamus. Also, cerebellar damage causes behavioral disturbance. Also, fMRI shows blood flow during purely cognitive tasks.