491
Unique features of the plant vacuolar sorting machinery
Diane C Bassham* and Natasha V Raikhel†
Multiple types of vacuoles can exist within the same plant cell,
and different vesicle-trafficking pathways transport proteins to
each of them. Recent work has identified proteins unique to
each vacuole type, and the transport pathways have begun to
be elucidated. Plant trafficking proteins are usually encoded by
small gene families, the different members of which have
distinct functions in the endomembrane system.
Addresses
Michigan State University - Department of Energy Plant Research
Laboratory, Michigan State University, East Lansing, Michigan
48824-1312, USA
*e-mail bassham@pilot.msu.edu
† e-mail nraikhel@pilot.msu.edu
Current Opinion in Cell Biology 2000, 12:491–495
0955-0674/00/$ — see front matter
© 2000 Elsevier Science Ltd. All rights reserved.
Abbreviations
CPY
carboxypeptidase Y
Cvt
cytoplasm-to-vacuole transport
PVC
prevacuolar compartment
SNARE SNAP receptor
TGN
trans-Golgi network
TIPs
tonoplast intrinsic proteins
Introduction
Proteins are transported to the plant vacuole by vesicle trafficking through the secretory pathway. As in other
organisms, most proteins are thought to enter the pathway at
the endoplasmic reticulum (ER) and are then transported
through the Golgi apparatus to the trans-Golgi network
(TGN). At the TGN, the pathways for secretion and transport to the vacuole diverge, with secretion being the default
pathway for soluble proteins [1]. A targeting signal within
the cargo protein is required for further transport to the vacuole. The signal can be one of three different types: a
cleavable amino-terminal or carboxy-terminal sorting signal
or a region within the mature protein. Transport involving
amino- and carboxy-terminal propeptides can be mechanistically distinguished using the inhibitor wortmannin [2],
providing evidence that in some cell types two separate
pathways exist for transport to the vacuole. Amino-terminal
sorting signals are recognized by the cargo receptor BP-80
(AtELP) [3–5], which causes the cargo to be packaged into
clathrin-coated vesicles [6] for transport to the prevacuolar
compartment (PVC) and ultimately the vacuole. Other proteins, typically vacuolar storage proteins containing a
carboxy-terminal sorting signal or a signal within the mature
protein, are transported to the vacuole in Golgi-derived
dense vesicles, rather than clathrin-coated vesicles [7], possibly via multivesicular bodies [8]. Certain storage proteins,
for example, in pumpkin seeds, may bypass the Golgi and
move to the vacuole in precursor-accumulating vesicles
(PAC) derived from the ER ([9], Figure 1).
In all eukaryotes studied, the fusion of a vesicle with its
target membrane during transport through the
endomembrane system involves proteins of the SNARE
(SNAP receptor) family. A v-SNARE on the vesicle
interacts with a t-SNARE on the target organelle, and
this interaction is required for membrane fusion. The
soluble factors NSF (N-ethylmaleimide sensitive factor)
and SNAP (soluble NSF attachment protein) are also
necessary, and the fusion reaction is regulated by a number of other proteins, including rab GTPases and
Sec1p-like proteins. In plants, members of each of these
families have been identified [1], and their specific functions are now being addressed.
In this review, we describe some of the advances made
over the past year in understanding targeting and transport
of proteins to the vacuole in plant cells. In particular, we
focus on those aspects that seem to be unique to plants and
on the differences that have recently emerged between
closely related proteins in yeast and plants.
Plant cells can contain multiple functionally
distinct vacuoles
One apparent oxymoron in plant cells is that vacuoles are
both storage and lytic organelles. In some cell types, storage proteins are stored in the vacuole for later degradation
and remobilization of their carbon and nitrogen. However,
the proteases involved in this degradation are also found in
vacuoles in these same cells.
Recent data have clarified this seemingly contradictory situation, as some plant cells, for example, those in seeds,
root tips and cotyledons, have two distinct vacuole types:
lytic vacuoles (equivalent to the yeast vacuole or mammalian lysosome) and protein storage vacuoles (Figure 1).
These can be distinguished by the presence of different
aquaporins in their limiting membranes (TIPs; tonoplast
intrinsic proteins) and lumenal contents [10–13]. It can be
inferred that different signals and pathways are required to
deliver proteins to the distinct vacuole types. A detailed
examination of these vacuoles using antibodies against different isoforms of TIP has now demonstrated that
different combinations of TIPs act as markers for vacuoles
with different contents and, therefore, presumably different functions [14•]. In addition, root tip cells were
identified containing three different vacuoles, each
marked by a distinct TIP isoform. Mechanisms must
therefore exist to generate and maintain these different
vacuoles within the same cell [14•]. The scarcity of marker proteins for the soluble protein content of the different
vacuole types complicates this analysis; the identification
of new markers for lytic and storage vacuoles is needed in
order to be able to draw more general conclusions about
the kinds of proteins present in each type of vacuole. In
492
Membranes and sorting
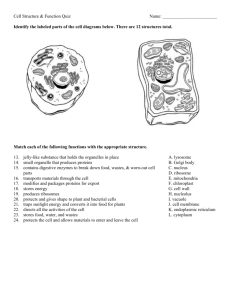
Figure 1
Protein-storage
vacuole
(a)
PA C
(b)
MVB
Fused
central
vacuole
DV
A hypothetical model showing pathways of
protein transport to the plant vacuole. In some
cell types, lytic vacuoles and protein-storage
vacuoles co-exist. After entry into the ER,
proteins are transported to the vacuole by one
of three major routes. (a) Precursoraccumulating vesicles (PAC) and (b) dense
vesicles (DV) transport storage proteins to the
storage vacuole, whereas clathrin-coated
vesicles (CCV) transport proteins to the lytic
vacuole. The relationship between the dense
vesicle and precursor-accumulating vesicle
pathways is still unclear, as is the role of
multivesicular bodies (MVB) in these
pathways. In mature cells, the lytic and protein
storage vacuoles fuse to produce a large
central vacuole.
TGN
(c)
Lytic
vacuole
Golgi
ER
CCV
Prevacuolar
compartment
Current Opinion in Cell Biology
many cells, the two types of vacuole are fused to form a
typical single large central vacuole containing both lytic
and storage vacuole marker proteins [15].
Golgi, and to fuse directly with the protein storage vacuole.
This pathway was proposed to play a role in the mobilization of storage proteins in germinating seeds [17•].
Developing pea cotyledons is one tissue where separate
lytic and protein storage vacuoles are evident, and this system has been used to begin the characterization of the
trafficking pathways to each type of vacuole. Hydrolases
are transported from the Golgi in clathrin-coated vesicles,
whereas storage proteins are transported in dense vesicles.
Purification of these two types of vesicles has allowed the
characterization and comparison of marker proteins within
them [16•]. Storage proteins and α-TIP, the TIP isoform
specific for protein storage vacuoles, were enriched in the
dense vesicle fraction, indicating that these proteins are
transported together to the storage vacuole. In contrast, the
cargo receptor BP-80 was found only in the clathrin-coated
vesicle fraction, not in the dense vesicles, consistent with a
role for BP-80 in transporting amino-terminal propeptidecontaining proteins to the lytic vacuole, whereas transport
to the storage vacuole may be independent of BP-80 [16•].
This pathway to the storage vacuole appears to be unique
to plants, and the components of the machinery that mediate it are not yet known.
Some proteins are localized differently in
Arabidopsis compared with yeast
The pathway for the transport of a protease to the protein
storage vacuole in mung bean cotyledons has been
described recently by Toyooka et al.[17•]. The protease
contains an ER retention signal that causes its accumulation as a proform in the ER. A large vesicle containing this
proform was shown to bud off from the ER, bypass the
Searches of the Arabidopsis thaliana EST and genomic
sequence databases revealed that Arabidopsis has numerous proteins that display similarity to those involved in
vesicle trafficking in other species [1]. When the localization of some endomembrane proteins in Arabidopsis and
yeast is compared, some differences become apparent
(Table 1). For example, AtVAM3 was identified as a cDNA
that is able to complement some phenotypes of a yeast
vam3 mutant [18]. Vam3p is a t-SNARE on the yeast vacuole that functions in multiple transport pathways to this
organelle [19]. The AtVAM3 protein was originally reported to reside on the tonoplast in the shoot apical meristem
[18]. Recently, however, it was shown that in roots and
leaves, AtVAM3 is found on the PVC and not the vacuole
[20]. The tonoplast localization may therefore be restricted
to specialized cell types in plants, and in most tissues
AtVAM3 is probably prevacuolar.
The Sec1p-like protein AtVPS45 also shows differences
when compared with its yeast homologue. AtVPS45 was
identified [21] by its sequence similarity to yeast Vps45p,
a protein required for transport to the vacuole via two distinct pathways: the CPY (carboxypeptidase Y) pathway
[22,23] and the Cvt (cytoplasm-to-vacuole transport) pathway [24]. It therefore seems to be a multi-functional
The plant vacuolar sorting machinery Bassham and Raikhel
Table 1
Summary of Arabidopsis proteins that may be involved in
vacuolar trafficking, their yeast equivalents and their locations
in plants, and the protein family they belong to.
Name
AtELP
AtPEP12
AtPLP
AtVAM3
AtTLG2a
AtTLG2b
AtVTI1a
AtVTI1b
AtVPS45
Yeast equivalent
Location
Protein family
none
Pep12p
Pep12p
Vam3p
Tlg2p
Tlg2p
Vti1p
Vti1p
Vps45p
TGN, PVC
PVC
Unknown
PVC
TGN
TGN
TGN, PVC
Unknown
TGN
Cargo receptor
t-SNARE
t-SNARE
t-SNARE
t-SNARE
t-SNARE
v-SNARE
v-SNARE
Sec1p
protein, interacting with the t-SNARE Pep12p at the PVC
in the CPY pathway and with the t-SNARE Tlg2p at the
late Golgi in the Cvt pathway. In contrast, AtVPS45 is
localized exclusively to the TGN in Arabidopsis roots and
interacts with two Tlg2p-like proteins: AtTLG2a and
AtTLGb. AtVPS45 does not interact with the Arabidopsis
Pep12p homologue AtPEP12, and AtVPS45 cannot be
detected at the PVC [25•]. The function of AtVPS45 thus
appears to have diverged from that of its yeast equivalent,
although, as yet, the precise function of AtVPS45 is not
known. The question remains as to whether plants have an
additional Sec1p homologue that interacts with AtPEP12
and functions at the PVC.
The difference in localization in plants and yeast is not
restricted to the vesicular trafficking machinery. The yeast
PVC plays a role in salt tolerance, for which the PVC Na+/H+
exchanger Nhx1 is required. Overexpression of the
Arabidopsis homologue AtNHX1 can suppress the nhx1 mutant
phenotype [26] and confer salt tolerance to Arabidopsis plants
[27]. However, AtNHX1 is found on the tonoplast, not the
PVC, in Arabidopsis, implicating the vacuole, rather than the
PVC, in salt tolerance via AtNHX1 in plants.
Arabidopsis has multiple isoforms of vesicle
trafficking proteins
In addition to differences in protein localization within the
endomembrane system, it is immediately apparent from
phylogenetic analyses [1] that, in many cases, for a single
yeast vesicle transport gene, multiple genes exist in
Arabidopsis (Table 1).
Yeast Pep12p is a syntaxin-like t-SNARE required for protein transport to the vacuole. It is localized to the PVC,
where it has been proposed to function in the fusion of
TGN-derived vesicles with this compartment [28]. An
Arabidopsis cDNA was identified by functional complementation of the yeast pep12 mutant encoding a protein
(called AtPEP12) with sequence similarity to yeast
Pep12p [29]. AtPEP12 resides on the Arabidopsis PVC
(and is in fact the first marker discovered for this compartment in plants [30]) and forms a complex with
493
characteristics of fusion complexes [31]. It was therefore
proposed that this protein is the Arabidopsis homologue of
yeast Pep12p. It is now apparent, however, that at least
two other genes encoding proteins that are highly related
to AtPEP12 exist in Arabidopsis. One of these genes is
AtVAM3 (described above), which encodes a t-SNARE of
the PVC [20]. Despite the ability of AtVAM3 to functionally replace Vam3p in yeast, its sequence is closely related
to AtPEP12 and yeast Pep12p and not yeast Vam3p. One
possibility is that vam3 mutant complementation may
reflect the mislocalization of the plant protein in yeast.
Interestingly, despite the sequence conservation, AtVAM3
is not able to complement the pep12 mutant, perhaps indicating that AtPEP12 and AtVAM3 do have distinct
functions. In addition, heterozygous Arabidopsis knockout
mutants have been isolated for the AtPEP12 and AtVAM3
genes, but we have been unable to generate plants
homozygous for either mutation, indicating that both
AtPEP12 and AtVAM3 are essential genes (AA Sanderfoot,
NV Raikhel, unpublished data).
The situation is further complicated by the discovery of a
third AtPEP12-like protein (AtPLP; [32]). Although
AtPLP has not yet been studied at the protein level, its
RNA can be detected in all tissues. Interestingly, in the
RLD ecotype of Arabidopsis, AtPLP encodes a typical syntaxin-like protein with a single carboxy-terminal
transmembrane domain, whereas in the Columbia ecotype
there is a frameshift in the genomic DNA resulting in a
stop codon, and the encoded protein thus lacks the transmembrane domain [32].
Similar observations have been made with other components of the vesicle-transport machinery in plants. For
example, the yeast TGN/endosomal t-SNARE Tlg2p also
has multiple homologues in Arabidopsis. At least two of
these, AtTLG2a and AtTLG2b, are expressed as protein
[25•]. Although both of these proteins interact with
AtVPS45, they are localized to distinct domains of the
TGN, indicating that they may have different functions
[25•]. The v-SNARE Vti1p, which is required for multiple
transport pathways in yeast [33,34], has at least two
expressed Arabidopsis isoforms, AtVTI1a and AtVTI1b,
that share approximately 60% amino acid identity.
AtVTI1a can substitute for Vti1p in yeast in the CPY pathway, whereas AtVTI1b substitutes for Vti1p in two
alternative pathways to the yeast vacuole: the ALP and Cvt
pathways [35•]. AtVTI1b, but not AtVTI1a, is a component of the complex containing AtVPS45 and AtTLG2a/b,
again suggesting that the isoforms represent a specialization of function, rather than simply redundancy [25•].
Finally, the plant vacuolar cargo receptor BP-80 (AtELP)
that recognizes proteins containing an amino-terminal vacuolar-sorting signal is a member of a small gene family in
Arabidopsis [4,5]. Although BP-80 (in pea; [36]) and AtELP
(in Arabidopsis; [37]) bind to amino-terminal signals, and
not to the carboxy-terminal signals tested, related proteins
494
Membranes and sorting
from other species appear to have somewhat different
specificities. Two pumpkin BP-80 homologues, PV72 and
PV82, can bind to internal and carboxy-terminal propeptide regions of the pumpkin 2S albumin [38], although the
Arabidopsis 2S albumin does not bind to BP-80 [36]. In
addition, immunoprecipitation experiments showed that a
tobacco BP-80-related protein complexes with a carboxyterminal propeptide-containing protease inhibitor protein
(Na-PI) in stigma tissue [39•]. The question therefore
remains open as to whether different BP-80 (AtELP) isoforms have tissue-specific expression or distinct substrate
specificities for different types of vacuolar-sorting signals.
The function of the individual isoforms of all of these proteins is now being investigated.
Conclusions
It is now clear that different vacuole types exist within the
same plant cell, and molecular markers are available for
each of these types. Efforts are now underway to characterize the differences in their protein content (both soluble
and membrane) and to elucidate the trafficking pathways
to each of them. Further analysis of the different vesicle
types carrying proteins to the vacuoles and the cargo found
in them will clarify their role in trafficking to a particular
type of vacuole. In addition, the receptor proteins responsible for the targeting of proteins with a carboxy-terminal
sorting signal remain to be identified (other than the few
possible exceptions described above) and this pathway
may be unique to plants.
the presence of distinct sorting machineries in tobacco cells.
J Cell Biol 1995, 130:1307-1318.
3.
Kirsch T, Paris N, Butler JM, Beevers L, Rogers JC: Purification and
initial characterization of a potential plant vacuolar targeting
receptor. Proc Natl Acad Sci USA 1994, 91:3403-3407.
4.
Ahmed SU, Bar-Peled M, Raikhel NV: Cloning and subcellular
localization of an Arabidopsis receptor-like protein that shares
common features with protein-sorting receptors of eukaryotic
cells. Plant Physiol 1997, 114:325-336.
5.
Paris N, Rogers SW, Jiang L, Kirsch T, Beevers L, Phillips TE, Rogers JC:
Molecular cloning and further characterization of a probable plant
vacuolar sorting receptor. Plant Physiol 1997, 115:29-39.
6.
Sanderfoot AA, Ahmed SU, Marty-Mazars D, Rapoport I, Kirchhausen T,
Marty F, Raikhel NV: A putative vacuolar cargo receptor partially
colocalizes with AtPEP12p on a prevacuolar compartment in
Arabidopsis roots. Proc Natl Acad Sci USA 1998, 95:9920-9925.
7.
Hohl I, Robinson DG, Chrispeels M, Hinz G: Transport of storage
proteins to the vacuole is mediated by vesicles without a clathrin
coat. J Cell Sci 1996, 109:2539-2550.
8.
Robinson DG, Bäumer M, Hinz G, Hohl I: Vesicle transfer of storage
proteins to the vacuole: the role of the Golgi apparatus and
multivesicular bodies. J Plant Physiol 1998, 152:659-667.
9.
Hara-Nishimura I, Shimada T, Hatano K, Takeuchi T, Nishimura M:
Transport of storage proteins to protein storage vacuoles is
mediated by large precursor-accumulating vesicles. Plant Cell
1998, 10:825-836.
10. Paris N, Stanley CM, Jones RL, Rogers JC: Plant cells contain two
functionally distinct vacuolar compartments. Cell 1996, 85:563-572.
11. Di Sansebastiano GP, Paris N, Marc-Martin S, Neuhaus JM: Specific
accumulation of GFP in a non-acidic vacuolar compartment via a
C-terminal propeptide-mediated sorting pathway. Plant J 1998,
15:449-457.
12. Jauh GY, Fischer AM, Grimes HD, Ryan CA, Rogers JC: δ-Tonoplast
intrinsic protein defines unique plant vacuole functions. Proc Natl
Acad Sci USA 1998, 95:12995-12999.
13. Swanson SJ, Bethke PC, Jones RL: Barley aleurone cells contain
two types of vacuoles: characterization of lytic organelles by use
of fluorescent probes. Plant Cell 1998, 10:685-698.
The complexity of plant vesicle trafficking and the differences between the plant and yeast endomembrane
systems have been highlighted recently by the identification of potential components for transport to the plant
vacuole. Unlike in yeast, most of these trafficking proteins
exist in small families, members of which are related in
sequence but probably have distinct functions. A major
challenge for the future is to define the function of these
proteins in the context of the whole plant, aided by the
availability of collections of Arabidopsis T-DNA and transposon knock-out mutants. The immunopurification of
specific organelles of the secretory pathway should also
yield valuable information about their cargo content, organization and function.
16. Hinz G, Hillmer S, Bäumer M, Hohl I: Vacuolar storage proteins and
•
the putative vacuolar sorting receptor BP-80 exit the Golgi
apparatus of developing pea cotyledons in different transport
vesicles. Plant Cell 1999, 11:1509-1524.
Dense vesicles were highly purified from developing pea cotyledons and
demonstrated to contain vacuolar storage proteins and α-TIP but not the
vacuolar cargo receptor BP-80, which was enriched in clathrin-coated vesicles. Storage proteins and α-TIP are thus delivered to protein storage vacuoles via the same pathway, independent of BP-80.
Acknowledgements
17.
•
This work was supported by research grants from the National Science
Foundation (MCB-9507030) and the Department of Energy (DE-FG0291ER-20021).
References and recommended reading
Papers of particular interest, published within the annual period of review,
have been highlighted as:
• of special interest
•• of outstanding interest
1.
Sanderfoot AA, Raikhel NV: The specificity of vesicle trafficking:
coat proteins and SNAREs. Plant Cell 1999, 11:629-641.
2.
Matsuoka K, Bassham DC, Raikhel NV, Nakamura K: Different
sensitivity to wortmannin of two vacuolar sorting signals indicates
14. Jauh GY, Phillips TE, Rogers JC: Tonoplast intrinsic protein
•
isoforms as markers for vacuolar functions. Plant Cell 1999,
11:1867-1882.
Different combinations of TIP isoforms are shown by immunofluorescence
analysis to mark vacuoles that are functionally distinct.
15. Schroeder MR, Borkhsenious ON, Matsuoka K, Nakamura K, Raikhel
NV: Colocalization of barley lectin and sporamin in vacuoles of
transgenic tobacco plants. Plant Physiol 1993, 101:451-458.
Toyooka K, Okamoto T, Minamikawa T: Mass transport of proform of
a KDEL-tailed cysteine proteinase (SH-EP) to protein storage
vacuoles by endoplasmic reticulum-derived vesicle is involved in
protein mobilization in germinating seeds. J Cell Biol 2000,
148:453-463.
The pathway for transport of the proteinase SH-EP containing an ER-retention signal to the vacuole was determined. The proteinase was packaged
into ER-derived large vesicles and delivered directly to the vacuole, bypassing the Golgi complex. The role of the protease in protein mobilization upon
seed germination is discussed.
18. Sato MH, Nakamura N, Ohsumi Y, Kouchi H, Kondo M,
Hara-Nishimura I, Nishimura M, Wada Y: The AtVAM3 encodes a
syntaxin-related molecule implicated in vacuolar assembly in
Arabidopsis thaliana. J Biol Chem 1997, 272:24530-24535.
19. Wada Y, Ohsumi Y, Hirata A: Vam3p, a new member of syntaxin
related protein, is required for vacuolar assembly in the yeast
Saccharomyces cerevisiae. J Cell Sci 1997, 110:1299-1306.
The plant vacuolar sorting machinery Bassham and Raikhel
495
20. Sanderfoot AA, Kovaleva V, Zheng H, Raikhel NV: The t-SNARE
AtVAM3p resides on the prevacuolar compartment in Arabidopsis
root cells. Plant Physiol 1999, 121:929-938.
31. Bassham DC, Raikhel NV: The pre-vacuolar t-SNARE AtPEP12p
forms a 20S complex that dissociates in the presence of ATP.
Plant J 1999, 19:599-603.
21. Bassham DC, Raikhel NV: An Arabidopsis Vps45p homolog
implicated in protein transport to the vacuole. Plant Physiol 1998,
117:407-415.
32. Zheng H, Bassham DC, Conceição AS, Raikhel NV: The syntaxin
family of proteins in Arabidopsis: a new syntaxin homologue
shows polymorphism between two ecotypes. J Exp Bot 1999,
50:915-924.
22. Cowles CR, Emr SD, Horazdovsky BF: Mutations in the VPS45
gene, a SEC1 homologue, result in vacuolar protein sorting
defects and accumulation of membrane vesicles. J Cell Sci 1994,
107: 3449-3459.
23. Piper RC, Whitters EA, Stevens TH: Yeast Vps45p is a Sec1p-like
protein required for consumption of vacuole-targeted, post-Golgi
transport vesicles. Eur J Cell Biol 1994, 65:305-318.
24. Abeliovich H, Darsow T, Emr SD: Cytoplasm to vacuole trafficking
of aminopeptidase I requires a t-SNARE-Sec1p complex
composed of Tlg2p and Vps45p. EMBO J 1999, 18:6005-6016.
25. Bassham DC, Sanderfoot AA, Kovaleva V, Zheng H, Raikhel NV:
•
AtVPS45 complex formation at the TGN. Mol Biol Cell 2000,
in press.
This paper demonstrates the presence of two protein complexes at the
Arabidopsis TGN, containing the Sec1p-like protein AtVPS45, the
v-SNARE AtVTI1b and either the t-SNARE at AtTLG2a or the closely related protein AtTLG2b. AtTLG2a and b are found in distinct subdomains of the
TGN, indicating that the two complexes may have different roles in protein
trafficking. Potential functions of these complexes are discussed.
26. Gaxiola RA, Rao R, Sherman A, Grisafi P, Alper SL, Fink GR: The
Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can
function in cation detoxification in yeast. Proc Natl Acad Sci USA
1999, 96:1480-1485.
27.
Apse MP, Aharon GS, Sneddon WA, Blumwald E: Salt tolerance
conferred by overexpression of a vacuolar Na+/H+ antiport in
Arabidopsis. Science 1999, 285:1256-1258.
28. Becherer KA, Rieder SE, Emr SD, Jones EW: Novel syntaxin
homolog, Pep12p, required for the sorting of lumenal hydrolases to
the lysosome-like vacuole in yeast. Mol Biol Cell 1996, 7:579-594.
29. Bassham DC, Gal S, Conceição AS, Raikhel NV: An Arabidopsis
syntaxin homologue isolated by functional complementation of a
yeast pep12 mutant. Proc Natl Acad Sci USA 1995, 92:7262-7266.
30. Conceição AS, Marty-Mazars D, Bassham DC, Sanderfoot AA,
Marty F, Raikhel NV: The syntaxin homolog AtPEP12p resides on a
late post-Golgi compartment in plants. Plant Cell 1997, 9:571-582.
33. Fischer von Mollard G, Nothwehr SF, Stevens TH: The yeast
v-SNARE Vti1p mediates two vesicle transport pathways through
interactions with the t-SNAREs Sed5p and Pep12p. J Cell Biol
1997, 137:1511-1524.
34. Fischer von Mollard G, Stevens TH: The Saccharomyces cerevisiae
v-SNARE Vti1p is required for multiple membrane transport
pathways to the vacuole. Mol Biol Cell 1999, 10:1719-1732.
35. Zheng H, Fischer von Mollard G, Kovaleva V, Stevens TH, Raikhel NV:
•
The plant v-SNARE AtVTI1a likely mediates vesicle transport from
the TGN to the prevacuole. Mol Biol Cell 1999, 10:2251-2264.
This paper describes the first characterization of two plant v-SNAREs,
AtVTI1a and b. When expressed in yeast mutants, the two Arabidopsis genes
appear to function in distinct pathways to the vacuole. Localization and interaction data also implicates AtVTI1a in transport to the vacuole in plants.
36. Kirsch T, Saalbach G, Raikhel NV, Beevers L: Interaction of a
potential vacuolar targeting receptor with amino- and carboxylterminal targeting determinants. Plant Physiol 1996, 111:469-474.
37.
Ahmed SU, Rojo E, Kovaleva V, Venkataraman S, Dombrowski JE,
Matsuoka K, Raikhel NV: The plant vacuolar receptor AtELP is
involved in transport of amino-terminal propeptide-containing
vacuolar proteins in Arabidopsis. J Cell Biol 2000, in press.
38. Shimada T, Kuroyanagi M, Nishimura M, Hara-Nishimura I: A pumpkin
72 kDa membrane protein of precursor-accumulating vesicles
has characteristics of a vacuolar sorting receptor. Plant Cell
Physiol 1997, 38:1414-1420.
39. Miller EA, Lee MCS, Anderson MA: Identification and
•
characterization of a prevacuolar compartment in stigmas of
Nicotiana alata. Plant Cell 1999, 11:1499-1508.
The authors used the precursor protein of proteinase inhibitors (Na-PI) in
tobacco stigmas as a marker to identify a prevacuolar compartment containing Na-PI with its carboxy-terminal targeting signal intact. Na-PI was
shown to associate with a BP-80-related protein in this compartment, indicating that in some cases, members of the BP-80/AtELP family may bind
carboxy-terminal signals as well as the well characterized binding to amino
terminal signals.