DRUG METABOLISM REVIEWS, 6(1), 1-50 (1977)
History of Drug Metabolism:
Discoveries of the Major
Pathways in the 19th Century*
A.CONTI and M. H. BICKEL
Medizinisch-chemisches Institut
University of Berne
Berne, Switzerland
I.
INTRODUCTION
2
II.
GLYCINE CONJUGATION
4
III.
OXIDATION
11
IV.
SULFURIC ACID CONJUGATION
20
V.
REDUCTION
25
VI.
GLUCURONIC ACID CONJUGATION
31
VII.
ACETIC ACID CONJUGATION
36
VIII.
IX.
GENERAL COMMENTS
SUMMARY-TABLES OF KEY DATES
A. Glycine Conjugation
B. Oxidation
C. Sulfuric Acid Conjugation
D. Reduction
39
43
43
43
44
44
*Based upon Dr. Conti's doctoral thesis (written in German) which received the Henry E. Sigerist award
by the Swiss Society of the His-tory of Medicine in 1975.
I
Copyright © 1977 by Marcel Dekker, Inc All Rights Reserved. Neither this work nor any part may be reproduced or
transmitted in any form or by any meafis, electronic or mechanical, including photocopying, microfilming, and
recording, or by any information storage and retrieval system, without permission in writing from the publisher.
2
CONTI AND BICKEL
E.
Glucuronic Acid Conjugation
44
F.
Acetic Acid Conjugation
45
References
45
I. INTRODUCTION
Biochemistry has a history of breathtaking successes and in addition has contributed immensely to the
progress of medicine during the past hundred years. Yet the history of biochemistry, unlike that of other
biomedical sciences, has attracted surprisingly little attention. One of the reasons is that biochemistry is
rather young compared with, say, anatomy or physiology. The very problem of biochemistry was
nonexistent before 1750, and only in the second half of the 19th century did it become a respectable
science. Although early biochemistry was strongly associated with organic chemistry, little help is
available from the well-covered history of chemistry, since chemists used to regard biochemistry either
as a chemical specialty of lower standing or else as a topic within the sphere of medicine. One of the
very few attempts at a comprehensive history of biochemistry is Geschichte der physiologischen Chemie
by Lieben [1], written in 1935. A new monumental work is being written by Florkin [2, 3]. Lieben's
book, although invaluable as a source of information, is an example of how difficult it is to portray the
development of a complex science like biochemistry without sacrificing either the chronology of its
authors or of its problems or even both. Thus the representation of a singled-out author or problem is a
much easier task which may still, and even in greater clarity, give the reader an idea of the development
and the spirit of early biochemistry.
This work is an attempt at representing the history of the metabolism of foreign compounds. The
problem of this topic emerged once scientists had realized that the living organism is a machinery
performing chemical reactions, and that synthetic compounds were becoming available in increasing
numbers. How, then, would the organism deal with compounds that were neither nutrients nor body constituents? This new line of biochemical research originated in the first half of the 19th century and indeed
became an important aspect to many of the pioneers of biochemistry. It is remarkable that all major
pathways of drug metabolism were discovered and verified during the 19th century: oxidation, reduction,
and the major conjugation reactions. Close interactions existed between these studies and general
biochemistry as well as neighboring disciplines like physiology and particularly pharmacology.
Buchheim, the very founder of modern pharmacology, as early as 1859 made the following statement:
HISTORYOF DRUG METABOLISM
3
In order to understand the actions of drugs it is an absolute necessity to have knowledge of the
transformations they undergo in the body. It is obvious that we must not judge drugs according to
the form and amount administered, but rather according to the form and amount which actually is
eliciting the action. [4]
What Buchheim had anticipated became one of the chief reasons for the pharmacologists' interest in drug
metabolism and for its amazing development in the course of the 20th century. A history of the early
stage of drug metabolism would be justified by the mere fact that many of the foremost old-day
biochemists, chemists, physiologists and pharmacologists were heavily involved, among them:
Woehler, Liebig, Naunyn, Nencki, Baumann, Jaffe, Bunge, Schmiedeberg. Most of their
activities took place in Germany, which was the major stage of early biochemistry.
The present review is restricted to the 19th century which not only contains the roots of drug metabolism
research but also sees it come to a certain completion. Emphasis is on the history of the discoveries of
the major pathways, with one section for each of the following reactions: glycine conjugation, oxidation,
sulfuric acid conjugation, reduction, and conjugations with glucuronic acid and with acetic acid. This
sequence was chosen for reasons of chronology and context of individual discoveries. Each section describes the history of the discovery from the first observations to the final proof. In addition to
developing the subject, which is based on the key publications, attempts have also been made at drawing
attention to the scientific background of the respective periods. All quotations have been translated from
the German originals. Thereby the language has been moderately modernized. A concluding section
deals with some aspects of the history of ideas and with practical problems of biochemical research in
the 19th century. Finally, tables of key dates and events were the only conceivable form of a summary
for this type of work.
The following references may be helpful to the reader who seeks access to the original literature on the
history of drug metabolism. Frankel [5] and particularly Heffter [6, 7] are the authors of the first and
most helpful reviews written shortly after 1900. Other authors have dealt with the topic in our time.
Williams [8], in his standard book Detoxication Mechanisms, gives a short survey of the history of his
subject matter. This coverage was later extended by Smith and Williams [9]. Some reflections have been
written by Butler [10], and Bickel [11], in a book on Nencki (1847-1901), has devoted one chapter to
drug metabolism.
4
CONTI AND BICKEL
II. GLYCINE CONJUGATION
The first compound to be investigated with respect to its fate in the body was benzoic acid. Being the
prototype for the study of glycine conjugation, it will also be the only example followed in this section.
The discovery of the biotransformation of benzoic acid is a long story of numerous, often contradictory,
observations dating back to the 18th century and gaining shape in the 1840s.
Benzoic acid was used therapeutically in the 18th century as a stimulant of the respiratory system and as
a tonic. The problem of the presence of benzoic acid in urine was discussed by Thompson [12] in 1802.
He was referring to Scheele who had evaporated urine, eliminated the salt component, and eventually
obtained "benzoic acid" by sublimation of the residue. According to Hallwachs [13], Roulle in 1773 was
the first to mention the presence of "benzoic acid" in cow's and later in camel's urine. These observations
were cited by Berzelius:
Rouelle found that from cow's urine which had been concentrated by evaporation, hydrochloric
acid would precipitate an acidic salt which he compared with benzoic acid. [14]
Fourcroy and Vauquelin [15] in 1799 also observed an acid in the urine of herbivorous animals which
they thought to be "benzoic acid." We purposely use quotation marks to indicate the likely incorrectness
of all these observations. Proust [16] as early as 1801 believed the acid found in urine to be similar to
rather than identical with benzoic acid.
The first researcher who set up an experiment to specifically investigate the fate of a foreign compound
was Woehler [17] in 1824. He gave a dog half a drachm of benzoic acid in its food and then isolated
from the urine an acid which crystallized in prismatic needles and had the general properties of benzoic
acid. According to Woehler this experiment proved that the compound absorbed in the gastrointestinal
tract reached the urine unchanged. Only in 1829 did Liebig [18] discover the compound he named
hippuric acid because he had isolated it from equine urine, and, in contrast to benzoic acid, it contained
nitrogen. Further investigations convinced Liebig that hippuric acid contained a benzoic acid moiety
which was bound to a nitrogenous compound. Retrospectively, it becomes evident that Woehler and his
predecessors were dealing with the then unknown hippuric acid which they mistakenly thought to be
benzoic acid. Erdmann [19], who also succeeded in isolating hippuric acid from equine urine, was
surprised to
HISTORY OF DRUG METABOLISM
5
find this acid mainly in saddle horses, whereas farm horses mainly excreted benzoic acid. This
observation was ascribed to different feeding, while Dumas [20] preferred the explanation that benzoic
acid would be formed as an artifact during the isolation procedure.
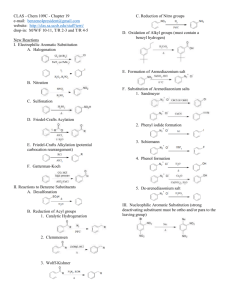
It was not until 1845 that the structure of hippuric acid was established when Dessaignes [21, 22] boiled
it with inorganic acids which made it split into its components, benzoic acid and glycine:
CONHCH2 COOH
COOH
+
Hippuric acid
Benzoic acid
NH2CH2COOH
Glycine
Dessaignes was also the first to perform a chemical synthesis of hippuric acid in vitro by
heating benzoic acid and glycine in a sealed tube for 12 hr at 1600C [22]. An empirical
formula could now be established beyond doubt. This exemplifies that in the 19th
century work on metabolic processes was highly dependent on progress made in organic
chemistry and particularly analytical methodology.
A decisive contribution to the problem of the fate of benzoic acid in the human body was made in 1841
by Ure [23], who knew that in the meantime hippuric acid had also been found in man [243. Administering benzoic acid to himself and to volunteers, Ure succeeded in isolating large quantities of
hippuric acid from the urine collections. He thus must be given the credit of being the first one to discover a biotransformation of a foreign compound. Since he also observed a concomitant decrease of uric
acid, he concluded that this nitrogen compound contributed to the formation of hippuric acid and he
therefore also suggested the use of benzoic acid for the treatment of gout.
After Liebig had discovered hippuric acid, Woehler [25], in a letter of 1830 to his friend, speculated on
the transformation of benzoic acid into hippuric acid during digestion. His pupil, Keller [26], felt
challenged by Ure's results, and with an experiment on himself was able to confirm that benzoic acid was
metabolized to hippuric acid. In contrast to Ure's finding, however, the excretion of uric acid was not
diminished by the administration of benzoic acid. Finally, Liebig [27] found that hippuric acid is also a
normal urinary product in humans consuming a mixed diet.
The first investigations focused upon the localization of glycine conjugation were carried out by Kuhne
and Hallwachs [28] in 1857. They must have been the first to study the localization of a biosynthetic
process and to use a foreign compound for the purpose. Less than a decade earlier, Woehler and Frerichs
[29] discussed the
6
CONTI AND BICKEL
problem of the localization of a metabolic process but dismissed it in view of the unsurmountable
difficulties. Kuhne and Hallwachs, however, very clearly stated their questions:
Is the ingested benzoic acid after its absorption transformed to hippuric acid by the action of the
blood, and does this process therefore occur in the whole vascular system? Or else, is benzoic acid
rather transformed by the action of bile, i.e., already in the intestinal lumen or within the hepatic
vessels?
[28]
The authors realized that the first question could only be answered by administering benzoic acid
intravenously. Another problem was the proper choice of the animal species to be used. Hallwachs and
Kuhne had observed that certain herbivorous species excreted hippuric acid without being treated with
benzoic acid. Thus only carnivorous species would be useful, and even with those the diet should be
strictly controlled to avoid vegetable components. In this case a serious source of error had been
recognized and avoided in time. This was not the rule in those days, and therefore false results and
conclusions, often to be perpetuated for decades, were frequent.
In a first series of experiments Kuhne and Hallwachs injected 20 dogs intravenously with benzoic
acid without, however, detecting more than traces of hippuric acid in the urine. They therefore switched
to test their “bile hypothesis" which had the following background. Strecker [30] had found that bile
contains the two bile acids, tauro cholic and glycocholic acid. Since no glycine could be isolated from
blood at that time, glycocholic acid was considered to be the only source of glycine in the body. In order
to eliminate this source, the bile ducts of dogs were cannulated according to the technique introduced by
Schwann [31] and Blondlot [32]. In their new series of experiments Kuhne and Hallwachs demonstrated
that animals with and without bile fistulae were transforming ingested benzoic acid to hippuric acid and
thus they proved that the presence of bile in the gut was not a prerequisite for the formation of hippuric
acid. Based on the presumption that bile acids are produced in the liver, they hypothesized that hippuric
acid is formed within the liver by using the glycine moiety of glycocholic acid. This hypothesis was to be
further tested by suppressing the liver function. Indeed, Kuhne and Hallwachs in 1857 seem to have been
the first to introduce such a technique by ligating the afferent and efferent liver vessels in cats. The
technique was adopted and improved by Eck in 1877 who introduced the classical portocaval shunt then
called Eck's fistula.
HISTORY OF DRUG METABOLISM
7
In the urine of cats with ligated hepatic blood vessels, Kuhne and Hallwachs were not able to
detect hippuric acid. Yet they did not draw the conclusion that the liver was the site of this synthesis
since they recognized that the portal ligation also blocked the reabsorption of bile acids from the gut.
Therefore, they injected cats intravenously with a mixture of benzoic acid and bile. The finding that
hippuric acid only was excreted in these animals led to the conclusion that the presence of glycocholic
acid in blood was essential to the conjugation of benzoic acid. At this stage the authors no longer took
into consideration that their results had been collected with two species, dogs and cats, which might have
influenced their reasoning in the light of the findings of Strecker [33] and Hoppe-Seyler [34] who
reported that dog bile had the extreme composition of 100% taurocholic acid and 0% glycocholic acid.
Why was this important information unknown to Kuhne and Hallwachs? Although there was no flood of
literature like in our times, not only the language but also the diffusion of scientific journals was
national. In many instances this situation prevented interaction between researchers.
In a third series of experiments, benzoic acid and a smaller amount of bile were injected into cats
and resulted in the excretion of benzoic acid in addition to hippuric acid. To Kuhne and Hallwachs this
finding was proof that the formation of hippuric acid was dependent on the presence of bile acids, a
condition met in the hepatic vascular system exclusively. Once they knew both the composition of
hippuric acid and the site of its formation, they felt ready for the venture of reproducing the biosynthesis
of this compound in vitro by incubation of benzoic acid with bile at 320 C. The authors must have been
rather perplexed by the total absence of hippuric acid:
It is hard to see what specific conditions exist in the organism. Their ignorance usually precludes
artificial simulations of physiological processes. [28]
This was the situation when 19 years later Bunge and Schmiedeberg [35] made another approach
toward the same problem. In the introduction of their publication they state that:
Many organic substances, when ingested, are hydrolytically split in the digestive tract: fat is split
into glycerine and fatty acids, starch into sugar. These processes are not puzzling to a chemist,
because they can be simulated in vitro. However, the situation is different with certain chemical
processes occurring with absorbed compounds within the
8
CONTI AND BICKEL
tissues... .The mechanism of the synthetic reactions (e.g., the reaction of benzoic acid and glycine
to form hippuric acid) is absolutely puzzling, because in vitro they can only be reproduced under
conditions which do not exist in the organism.
[35]
Thus biochemical reactions were subdivided into hydrolyses, which were easy to simulate and to
understand, and into still mysterious biosyntheses. The very discrepancy between the fast-growing art of
the synthetic chemists on one hand and the total lack of knowledge concerning biosyntheses on the other,
was a stimulus to investigate the latter, whereby the glycine conjugation of a foreign compound became
an important model. The conflict of the laws of chemistry existing in the test tube and in living
organisms was, in fact, one of the great challenges for the early biochemists.
More than their predecessors Kuhne and Hallwachs [28], who indicated the liver vessels as the
site of hippuric acid formation, and Meissner and Shepard [36], for whom it was the kidney, Bunge and
Schmiedeberg were aware of the difficulties and requirements of their project. The experiments were
carefully planned, and the chemical methodology was given fall attention:
The fact that previous investigations on hippuric acid formation in the body have not produced
unequivocal results is mainly due to the use of insufficient methods for the detection of hippuric
acid in tissues and body fluids. It was our prime task to develop a method which was absolutely
certain to detect even minute amounts of hippuric acid and which was not disturbed by the other
constituents of the animal body.
[35]
Again, the localization of the process was their first target. After injections of both benzoic acid and
glycine into operated dogs, hippuric acid was detected in the blood of an animal with ligated hepatic vessels, but not in another one whose renal vessels were ligated. This was a first hint that the kidneys might
be involved in this biosynthesis. In intact animals the blood contained both benzoic and hippuric acid,
whereas the urine contained exclusively hippuric acid. Dogs with ligated ureters showed higher blood
concentrations of hippuric acid. Bunge and Schmiedeberg were not yet convinced that the kidney was
the sole site of hippuric acid synthesis. In addition, they realized that their ligation of hepatic vessels
resulted in serious disturbances of the hemodynamic situation. They therefore developed a technique
HISTORY OF DRUG METABOLISM
9
of total hepatectomy which became famous 10 years later under the authorship of Minkowski. The first
animals to be hepatectomized by Bunge and Schmiedeberg were frogs. Not only were they still able to
produce hippuric acid when benzoic acid was injected with glycine, but they also did so when benzoic
acid was injected alone. This observation disproved the theory that the liver was the origin of the glycine
moiety of hippuric acid. Still, Bunge and Schmiedeberg were not happy with their unphysiological
technique of total organ extirpation. They therefore adopted the technique of artificial perfusion of
isolated organs which had been developed by Ludwig and Schmidt [37] in 1868. In their first experiment
Bunge and Schmiedeberg perfused a kidney for 8 hr with blood containing benzoic acid and glycine.
Hippuric acid appeared in both blood and urine. This success led to the new question, namely,
whether glycine which was added to the blood participates in hippuric acid formation, or whether
the glycine moiety of hippuric acid is formed from some constituents of blood or kidneys and
combines with benzoic acid in statu nascendi.
[35]
This problem had just been investigated in 1875 by Spengel [38] in Nencki's laboratory who gave
an intact dog 3 g of benzoic acid in food and found equal amounts of benzoic and hippuric acid in the 24hr urine. If, however, the same dog was given 5 g of benzoic acid together with the same amount of
glycine, no unconjugated benzoic acid appeared in the urine. This demonstrated that the dog was able to
use exogenous glycine for the conjugation of benzoic acid. The same observation was made in man by
Hoffmann [39] in 1877. On the other hand, Bunge and Schmiedeberg were wondering about being
unable to verify these results, thereby overlooking that their experimental conditions (i.v. and 3.5 hr
urine instead of p.o. and 24 hr) were different in essential details. It was another advantage of the
perfusion of isolated organs that this technique eliminated pharmacokinetic factors which were much
neglected at that time. Further experiments showed that after several hours of perfusion of a kidney with
drug-free blood no hippuric acid was formed, whereas the addition of benzoic acid in-to the perfusion
system of the other kidney of the same dog led to the formation of hippuric acid. Its amount, however,
was much lower than with concomitant addition of glycine. Again, new and important questions came
up. Was the biosynthetic ability due to the organ as such, to its cells, or even to chemical constituents of
the cells? By perfusing kidneys 5 hr post-mortem or after a 48-hr storage in an ice
10
CONTI AND BICKEL
chest, the authors were still able to detect the formation of hippuric acid. On the other hand, fresh calf
kidneys, extensively homogenized in a mortar and incubated with benzoic acid and glycine, did not yield
hippuric acid.
This result was confirmed by Koch [40] in Pfluger's laboratory. If, however, he only mildly
minced the kidneys with a meat grinder, hippuric acid was formed. The more Koch homogenized the
tissue in his grinder, the less hippuric acid did he obtain. He concluded that
these experiments demonstrate that hippuric acid is synthesized by the vital functions of the
individual intact cells when exposed to benzoate and glycine in oxygenated blood. [40]
What, then, was the role of oxygen in glycine conjugation? Bunge and Schniiedeberg realized that this
problem was well suited for study with their perfusion technique. When they used saline or plasma
instead of whole blood as a perfusion medium, hippuric acid was detectable in traces only. They
concluded that red cells play an important role in glycine conjugation, but that very likely they serve as
mere carriers of oxygen.
Hoffmann [39], also a member of Schmiedeberg's team, modified that experiment by using whole
blood whose oxygen had been displaced by carbon monoxide. No hippuric acid was formed by the
perfused kidney. But then this could have been due to lack of oxygen as well as to a toxic effect of
carbon monoxide. Another approach tried by Hoffmann was the use of quinine instead of CO. Why
quinine? Studies carried out by Binz [41, 42] and by Scharrenbroich [43] had shown that low
concentrations of quinine block the motility of white blood cells and protozoa. Since all vital functions
seemed to disappear, Hoffmann concluded that quinine had a specific cytotoxic effect. His use of quinine
is of particular interest, since this may have been the first attempt to influence the metabolism of a drug
by a second or "tool" drug. Hoffmann indeed was successful in demonstrating that with increasing
amounts of added quinine the amount of hippuric acid formed was decreasing. He thereby also indicated
the possibility of metabolic interactions between drugs, one of the problems to be rediscovered a hundred
years later'.
Glycine conjugations of compounds other than benzoic acid are mentioned in the first half of the
following section.
HISTORY OF DRUG METABOLISM
11
III. OXIDATION
Ure [44] and Erdmann and Marchand [45, 46] in 1842 were the first to indicate that acids other
than benzoic acid are also excreted as hippuric acid. Being interested in the fate of cinnamic acid, the
latter authors gave 5 to 6 g of the compound in syrup to six volunteers. The urine of all volunteers then
disclosed a compound which was likely to be hippuric acid, although the glycine conjugate of cinnamic
acid could not be excluded [47]. This was one of the many instances in which early biochemists
approached a problem without sound knowledge of the underlying organic chemistry and without
adequate analytical methodology. Even though Erdmann and Marchand were well aware of their
equivocal result, they were eager to draw attention to the possibility of an oxidation-conjugation
sequence in the metabolism of a foreign compound according to the (modern) reaction scheme:
H
C C COOH
H
Cinnamic acid
O
COOH
Benzoic acid
Gly
Hippuric acid
This hypothesis of oxidative processes in drug metabolism was confirmed in 1848 by Wohler and
Frerichs [29] who administered cinnamic acid to dogs and indeed detected hippuric acid as the urinary
product. The same product was also isolated in considerable amount from the urine of dogs and rabbits
which were fed oil of bitter almonds (benzaldehyde). Based on their sound analytical methodology,
Wohler and Frerichs must therefore be given credit of having discovered the metabolic oxidation of
benzaldehyde to benzoic acid and with having formulated the reaction sequence of oxidation and
subsequent glycine conjugation. In the same work [29] these authors also made a noteworthy statement
on the importance of metabolic studies with foreign compounds for physiology and biochemistry at
large:
The transformations which substances undergo in the animal body between their ingestion and
excretion in the urine, are of considerable interest to physiology and biochemistry, because we
thereby may expect to gain insight into the chemical forces which are acting in the organism and
particularly in blood, and which are responsible for the metabolic processes. In particular this is
the case when compounds are
12
CONTI AND BICKEL
studied whose chemical constitution and properties are well known and whose metabolic
transformation thus allows conclusions with respect to the cause giving rise to metabolism.
Obviously, in order to obtain general results along these lines, extensive and time-consuming
series of experiments will be required and particular difficulties will have to be overcome. [29]
A new impulse came from studies on the volatile constituents of urine, carried out in 1851 by
Staedeler [48], who isolated a compound he believed to be phenol. He was surprised to find a compound
known as a poison in the urine of healthy humans and animals. Based on a report by Schlieper [49], who
had considered phenol an oxidation product, Staedeler speculated on the formation of phenol by metabolic oxidation of aromatic precursors. Although Staedeler's observation was to be disproved, his
hypothesis became one of the starting points for the famous investigations of Schultzen and Naunyn [50]
in 1867. These authors were interested in the fate of therapeutically used hydrocarhons such as benzene,
which was prescribed as an intestinal disinfectant. They first developed a more sensitive method for the
detection of phenol by adding ferric chloride to the distillate of urine, which resulted in a dark violet
color. Whereas this test was negative with urine of untreated animals and humans, phenol was detected
in patients, normal humans, and dogs treated with benzene. These results led Schultzen and Naunyn to
the following conclusion:
Thus, it seems that benzene during its passage through the organism undergoes a simple oxidation
which leads to the formation of phenol. If this result can be adequately verified, it would be of
great interest since it has never been possible so far to obtain simple derivatives by direct oxidation
of benzene. [50]
As has been mentioned, the biosynthesis of hippuric acid surprised its investigators because the
reaction in vitro would progress under extreme conditions only. Another, even more puzzling situation
was now introduced by the hydroxylation of benzene, a reaction unheard of in organic chemistry. The
latest attempts at chemical oxidation of benzene by Carius [51] resulted in the formation of formic acid,
benzoic acid, and phthialic acid, but not phenol.
Another problem in the investigation of Schultzen and Naunyn remained unsolved: Only a small
fraction of the ingested benzene was excreted as phenol. What, then, happened to the remainder? They
HISTORY OF DRUG METABOLISM
13
speculated that either phenol was further oxidized in the body or else that benzene was also excreted by
the lungs. Both hypotheses would later be proved correct.
Having discovered the metabolic hydroxylation of benzene, Schultzen and Naunyn were now
eager to study and compare the fate of benzene derivatives. Toluene, the first compound of a series, was
found to be oxidized in the sidechain rather than in the aromatic nucleus, thus forming benzoic acid
which was then conjugated with glycine:
CH3
COOH
Toluene
Benzoic acid
COGly
Hippuric acid
This finding, which was soon confirmed by Nencki [52], demonstrated still another foreign compound to
be ultimately excreted as hippuric acid. Many further instances of glycine conjugations were soon to be
discovered. Another model compound used by Schultzen and Naunyn in their systematic investigation
was o-xylene. This compound was recovered in the urine of dogs and humans as an acid, identical to
toluric acid, which had been described by Hofmann [53] and Kraut [54, 55] as the urinary glycine
conjugation product of toluic acid.
o-Xylene
CH3
CH3
CH3
CH3
COOH
COGly
o-Tolulc acid
Toluric acid
Again, Schultzen and Naunyn had discovered an oxidation-conjugation sequence. In addition, they
stressed the fact that only one of the two sidechains attached to the benzene ring was oxidized and
conjugated.
Working along the same lines, Nencki [52] (Fig. 1) was wondering which of two sidechains of
different sizes would be metaholically attacked. He cited his consideration of a recent chemical study of
Fittig and K6nig [56] who showed that only the longer of two side-chains was oxidized by hoiling nitric
acid. This chemical finding gave Nencki the idea to use p-cymene as a substrate. In 1872 Neneki and
Ziegler [57] were able to detect cumic acid in the urine of dogs and humans given 3-g doses of the
hydrocarhon. Thus, contrary to nitric acid, biological oxidation attacked the smaller of the two sidechains. Apparently the acid metabolite was not conjugated with glycine but instead was excreted
unchanged, an observation which had also been made by Hofmann [53], Kraut [55], and Buchheim [58]
who found cumic acid to be excreted unchanged. Later on, however,
14
CONTI AND BICKEL
FIG. 1. Marceli Nencki, 1847-1901. Berlin, Berne, St. Petersburg.
Jacobsen [59], after giving a dog 11 g of cymene, was able to detect cuminuric acid, the glycine
conjugate:
H3C
H3C
p-Cymene
CH3
H3C
H3C
COOH
H3C
COGly
H3C
Cumic acid
Cuminuric acid
HISTORY OF DRUG METABOLISM
15
As a further step, Nencki [60] then studied the fate of a benzenoid hydrocarbon with three sidechains,
mesitylene, which again was found to be oxidized in only one of the sidechains:
H3 C
H3 C
H3 C
CH3
H3C
COGly
COOH
H3 C
H3C
Mesitylene
Mesitylene
carbexylic acid
Mesitylenuric
acid
Another study in this field was carried out by Schultzen and Graebe [61] who felt disturbed about
the conflicting reports concerning glycine conjugation of aromatic acids. Thus Bertagnini [62] ingested
small amounts of nitrobenzoic acid and, noticing no adverse effects, increased the dose to a total of 6 g,
after which he was able to detect nitrohippuric acid in his urine. This was confirmed in the dog by Jaffe
[63]. In a later study Bertagnini [64] reported that salicylic acid given to humans is excreted partly
unchanged and partly as its glycine conjugate, salicyluric acid. In contrast, Wohler and Frerichs [29],
working with dogs, found unchanged salicylic acid only. Bertagnini [64] also found that anisic acid did
not undergo glycine conjugation. Similarly, Beilstein and Schiun [65] reported chlorobenzoic acid, and
Berg [66] coumaric acid, to be excreted unchanged. All these results prevented Schultzen and Graebe
from discerning a rule for the glycine conjugation of aromatic acids. At the same time these authors also
had their doubts about the methodology used by some of their predecessors and therefore decided to
repeat certain experiments. They selected cinnamic acid, chlorobenzoic acid, and anisic acid, which they
ingested. Their urine collections were found to contain hippuric acid, chlorohippuric acid, and anisuric
acid, respectively.
H3CO
COOH
Anisic acid
H3 CO
COGly
Anisuric acid
16
CONTI AND BICKEL
From their experiments, Schultzen and Graebe drew the conclusions that carhoxyl groups attached to the
aromatic nucleus are directly conjugated with glycine, whereas in aryl aliphatic acids the sidechain is
first oxidized and only subsequently does the newly formed carboxyl group undergo glycine conjugation.
Thus all aromatic acids are eventually transformed to glycine conjugates. Although the series of
compounds investigated at the time was too small to allow the establishment of such a rule, glycine
conjugation had definitely been shown to be a biochemical principle rather than an isolated reaction of
benzoic acid. A final experiment carried out by Schultzen and Graebe was aimed at elucidating the
metaholic fate of the aromatic dicarboxylic acid, phthalic acid, since doubly glycine-conjugated acids
would have been a novelty. Unfortunately, the small amounts of isolated urinary metabolite precluded
elementary analysis and establishment of the empirical formula.
Another unsuccessful attempt at identifying the metabolite of phthalic acid was made by Nencki,
who was doing his thesis work as a medical student under Schultzen's guidance. In this tbesis of 1870,
entitled “Oxidations of Aromatic Compounds in the Animal Body” [52], Nencki not only formulated the
rules of oxidation and glycine conjugation, which he himself was to enrich and improve by his lifework,
but he also characterized the potential of drug metabolism studies:
There can be no doubt that chemical groups and their homologues not only show an analogous
behavior toward certain chemical reagents, but also, if within limits, toward the animal organism.
By studying the metabolic fate of chemical substances, one will on one hand be able to establish
laws allowing predictions on the fate of new compounds, and on the other hand gain increasing
insight into the organism as a “chemical agent”. Of course, sound knowledge of the chemical
structure of compounds is a prerequisite to the study of their metabolic transformations.
[52]
A decade after the discovery of the metabolic oxidation of benzene to phenol by Schultzen and
Naunyn, Munk [67] expressed renewed surprise about this reaction which could not be performed in the
test tube even using the strongest oxidants. Two reasons caused him to repeat these experiments: first his
conviction that this was a question of fundamental importance in the light of biological oxidation, and
second the availability of an improved method for the determination of phenol
HISTORY OF DRUG METABOLISM
17
as tribromophenol, introduced by Landolt [68]. Munk and volunteers ingested 20 drops of benzene.
Although the experiment was performed according to a carefully prepared protocol, Munk was not able
to detect phenol in the urine. Only in the light of sulfuric acid conjugation could this dilemma be solved
in the same year. However, Munk correctly interpreted part of the fate of benzene in his comment:
Part of the benzene is excreted unchanged as a gas from the stomach. During the first 8 hr after
ingestion we were frequently annoyed by hiccups which had the exquisite smell and taste of coal
tar and therefore were rich in benzene vapors. We do not know whether benzene is also excreted
by the lungs, as suggested by Schultzen and Naunyn, nor, most important, what happened to the
remaining benzene. [67]
So far, studies on the metabolism of foreign compounds were motivated by scientific curiosity.
Nencki and Sieber [69-71] in the early 1880s may have been the first ones to think of clinical applications. Typically, they associated a particular metabolic disorder with a chemical model system. The
metabolic disorder in diabetes was characterized by the carbohydrates being excreted without having undergone combustion. Thus the underlying defect was believed to be an insufficient oxidation capacity of
the body. At the same time Nencki and Sieber were dealing with a new and fascinating chemical system,
able to oxidize glucose to lactate by simply incubating the substrate at 370C with strong alkali in the
presence of oxygen for 24 hr. Working with this so-called Radziszewski system, they made the important
observation that benzene was oxidized to phenol and, furthermore, that this oxidation was suppressed in
the presence of glucose. Thus a competition of the two substrates for oxygen was assumed, and this principle was then applied to the organism. If, in the body, glucose and benzene would compete for a
common oxidation system, then the oxidation capacity of the organism could be tested by measuring the
phenol formed from a dose of benzene. In contrast to biological substrates, benzene is not degraded to
C02 and H20, and its oxidation product, phenol, is still an aromatic compound which can easily be
detected and determined. Actually, Nencki in his extensive studies on the fate of aromatic compounds
made use of the stable benzene ring as a label, much as isotopically labeled molecules were used almost
a century later.
In order to use the “benzene test” to measure oxidation capacity under pathological conditions in
patients, the constancy and reproducibility of phenol formation was carefully studied in healthy rabbits,
18
CONTI AND BICKEL
dogs, and humans. Indeed, in all three species standard doses of benzene led to comparable amounts of
free and conjugated urinary phenol, independent of diuresis and nutritional state. Nencki and Sieber were
now convinced that the benzene test was superior to the then used determination of urinary lactate as an
indicator of the “ intensity of vital combustion.”
The first patient on whom the benzene test was performed was a diabetic woman. She was given
an oral dose of 6 g of benzene, and urine was collected for 4 days and analyzed for the presence of
phenol. The fact that its amount was comparable with that of healthy persons was interpreted as an
indication that diabetes was not due to a deficient oxidation capacity of the organism. In a patient with
splenic leukemia, the benzene test resulted in decreased amounts of phenol. Since, however, in this case
even under lactate treatment the patient’s urine remained free of lactate, Nencki and Sieber were inclined
to assume hypoxia to be due to low red cell counts rather than to an impaired oxidation capacity of the
tissues. On the other hand, Schultzen and Riess [72] and Quincke [73] had reported that patients with
phosphorus poisoning excreted lactate but had normal hemoglobin values. Thus this kind of poisoning
was selected as a model of likely oxidation deficiency by Nencki and Sieber. For obvious reasons the
benzene tests were now performed on animals. They clearly showed a decrease of phenol excretion
which corresponded to the degree and duration of phosphorus poisoning.
Drug metabolism studies were not only carried into the realms of diagnostics and etiology, but
also played a role in the long and tedious process of understanding the mechanism of biological
oxidation. Up to the 187Os the oxidation of nutrients and body constituents was believed to take place in
the blood. One of the first attempts at explaining why and how oxygen in blood was able to oxidize
otherwise stable substrates was based on Schonbein's discovery of ozone and its oxidative power in
1840. In 1862 Schmidt [74] postulated the presence of ozone in blood to explain the oxidations. This
hypothesis was supported by Schonbein [75] on the grounds that ozone is formed by atmospheric
electricity and thus must be present in air. Another hypothesis was expressed by Clausius [76] who
postulated oxygen splitting by some biological process, thus forming a mixture of 02, 03, and easily
available Ol as the actual oxidant. A totally different explanation was offered in
Traube 's work Theory of Enzyme Effects [77] in 1858. This author postulated body constituents being
able to bind and transfer oxygen much like catalysts. As an analogy he mentioned alcohol which is
oxidized by oxygen only in the presence of highly dispersed platinum as an apparent oxygen carrier. The
time was not
HISTORY OF DRUG METABOLISM
19
ready for Traube's remarkably modern theory. On the other hand, it became clear by 1870 that ozone was
not a biological constituent. What remained of the ozone theory was the concept of nascent oxygen. Thus
Binz [78] differentiated between 02, which is taken up by the lungs and transported by the blood to the
tissues on the one hand, and O1 which is formed in the tissues from 02. Binz [78] and Pfluger [79] were
the ones who in 1872 showed that biological oxidation takes place in the solid tissues rather than in
blood. Still another oxidation theory was postulated by the eminent and most influential biochemist,
Hoppe-Seyler [80] in 1877. His theory originated from observations in the fields of fermentation and
putrefaction, and also from the discovery that benzene could be oxidized to phenol by oxygen in the
presence of hydrogen adsorbed to palladium. According to Hoppe-Seyler's explanation, nascent
hydrogen of the palladium surface would react with one oxygen atom of 02, thereby forming nascent
oxygen which in turn would oxidize benzene. Fascinated by the oxidation of benzene to phenol in the
body, Hoppe-Seyler and his students went so far as to postulate the formation of nascent hydrogen in the
tissues as the actual source of nascent oxygen.
This hydrogen theory was never accepted by Nencki. On the other hand, he and his colleague
Kaufmann [81] were still intrigued by the similarities between oxidations in the animal body and by
ozone, even though they accepted the view that ozone was not a physiological agent. Indeed, Nencki and
Giacosa [82] were able to obtain phenol by blowing ozonized oxygen through benzene in the absence of
any catalyst. At the same time Radziszewski [83] introduced the already mentioned model system which
oxidized substrates in the presence of oxygen and strong alkali. Studying the question of how molecular
oxygen is split in the tissues, and experimenting with the Radziszewski system, Nencki and Sieber [69,
70] investigated the possible role of alkali in living cells, which they dismissed. Rather, they developed
Traube's [77] hypothesis and also indicated the interior of tissue cells as the site of biological oxidation:
Many experiments have shown that protein of the living protoplasm is able to strongly absorb
oxygen. The reason for this affinity must therefore be sought in the molecular architecture of that
protein.... It must be concluded that the oxidizing function of cells consists in their formation of
easily oxidizable, strongly reducing substances, the major one being the protoplasm itself, which is
made up of a labile protein molecule This protein molecule must be oxidizable by molecular
oxygen much like cuprous oxide or
20
CONTI AND BICKEL
benzaldehyde and at the same time must form atomic oxygen which in turn oxidizes cellular
substrates which are not oxidizible by molecular oxygen. As can be seen, the questions to be
answered will first of all depend on progress in chemistry, such as the discovery of new
compounds which are oxidizable by molecular oxygen, the understanding of the reactions
involved, the study of the composition of the protein molecule with respect to its fragments of
cleavage. Until we shall be able to represent the mechanism of oxidation in an animal cell by a
simple chemical equation, all these difficult questions will have to be answered by chemical
experimentation. [70]
Thus Nencki in 1880, at a time when terms like “ protein” or “ enzyme” were extremely vague,
anticipated the principle of respiration chain enzymes. Obviously, the oxidation of benzene and other
foreign compounds had helped a great deal.
IV. SULFURIC ACID CONJUGATION
As has been mentioned before, Munk [67] in 1876 speculated on the fate of the major part of
ingested benzene that did not appear as urinary phenol. He quoted Buliginsky [84] who in 1867 had
reported a compound in bovine and equine urine which, upon heating with dilute acids, was split and
formed phenol. This observation had been confirmed in the meantime by Hoppe-Seyler [85]. Apparently
these authors were not aware of the compound, phenylsulfuric acid, which had been synthesized by
Staedeler [86] from phenol and sulfur trioxide in 1867. Since phenol in urine was only obtained after
acid hydrolysis, Munk [67] assumed this compound to be bound to an unknown component, and he
named the hypothetical compound “ phenol-forming substance.” After ingesting varying amounts of
benzene, Munk was able to detect a direct relationship between benzene dose and phenol-forming
substance excreted. He thereby confirmed his hypothesis that a considerable part of benzene is
transformed into phenol which is bound to an unknown constituent and that this indicates another type of
conjugation in addition to the well-known glycine conjugation.
Munk's phenol-forming substance was not the only example of an unidentified hydrolyzable
constituent of urine. An indigo-forming substance (indican) was found in urine by various authors, and
Baumann [87] (Fig. 2) in 1876 detected a catechol-forming substance in the urine of horses and humans.
In the same year he performed an
HISTORY OF DRUG METABOLISM
21
FIG. 2. Eugen Baumann, 1846-1896. Tubingen, Strassburg,
Berlin, Freiburg.
analysis of the components of the indigo-forming substance, showing that:
if purified indigo-forming substance is split with acetic or hydrochloric acid, one always obtains a
certain amount of sulfuric acid which had not been present in the solution before.... This substance
... therefore seems to be a conjugated sulfuric acid whose carbon-containing atomic complexes
have not yet been identified. [88]
Further investigations led Baumann to the conclusion that sulfuric acid conjugates are regular
constituents of human, canine, and particularly equine urine. In the same year and still under the
influence of his teacher, Hoppe-Seyler, Baumann tried to connect his discovery of sulfuric acid
conjugation to Munk's reported phenol-forming
22
CONTI AND BICKEL
substance [88]. From horses' urine he isolated a considerable amount of phenol-forming substance which
he was then able to split into phenol and sulfuric acid by acid hydrolysis [89]. Thereupon he verified his
finding by administering phenol to volunteers. While their urine did not contain the unchanged
compound, large amounts of phenol and sulfuric acid were obtained upon hydrolysis. Sulfuric acid
conjugation of catechol was also observed. Thus conjugation with sulfuric acid began to emerge as a
biochemical principle.
Baumann also succeeded in identifying the chemical structure of the phenol-forming substance
and of sulfuric acid conjugates in general. Elementary analyses yielded the
empirical formula C6H5KS04. He realized that this formula could apply either to one of the three isomeric phenol-sulfonic acids (HO)C6H4SO2OK or else to phenylsulfuric acid C6H5SO4K. Each of the
sulfonic acid structures would contain
a phenolic hydroxyl group which would react with alkyl halide to form an ether. Since this could not be
achieved, Baumann concluded that the conjugate was an ether of phenol and sulfuric acid. He then once
more went back to the metabolism of benzene, this time by measuring not only urinary phenol but also
sulfuric acid [90]. This resulted in the remarkable observation that the ingestion of benzene led to a 10to 15-fold increase of total urinary sulfuric acid as determined after hydrolysis.
Baumann [91] also succeeded in identifying “ taurinic acid”, which had been found in urine and
described by Staedeler [48], as the sulfuric acid conjugate of p-cresol. This finding was confirmed in
1877 by Baumam and Herter [92] in a study involving a series of phenols, phenol homologs, and
aromatic compounds. Some, but not all, of the latter were metabolized and excreted as sulfuric acid
conjugates. Among them was indole which was hydroxylated to indoxyl and excreted as indoxyl sulfate
(indican), the indigo-forming substance mentioned above. Of particular interest was the fate of aniline,
which Woehler and Frerichs [29] in 1848 could not find in the urine after oral administration. This had
been confirmed in the meantime by Schuchardt [93] and Sonnenkalb [94]. Baumann and Herter [92]
found aniline to be hydroxylated to p-aminophenol and conjugated to p-aminophenyl sulfate, which was
soon to be confirmed by Schmiedeberg [95].
H2N
Aniline
H2N
OH
p-Aminophenol
H2N
OSO3 H
p-Aminophenyl sulfate
HISTORY OF DRUG METABOLISM
23
The studies of Baumann and Herter made it clear that, as with glycine conjugation, sulfuric acid
conjugation was often preceded by oxidation of the drug. In contrast to these authors, however,
Schmiedeberg [96] in 1881 postulated an inverse sequence:
+
O
+
H2SO4
OSO3H
+ H2O
OH
+ H2SO4
The question was settled by an elegant argumentation in a paper of Nencki and Sieber [71]. In
phosphorus poisoning the oxidation of benzene to phenol was blocked, whereas the sulfuric acid
conjugation of administered phenol was the same as before poisoning. This proved the oxidationconjugation sequence to be the correct one.
Another problem studied by Baumam and Herter [92] was the fate of hydroxybenzoic acids.
These compounds were promising models for the study of conjugation processes since they contained
both an acidic group, likely to conjugate with glycine, and a phenolic group, likely to conjugate with
sulfuric acid. In agreement with Bertagnini [64], the ortho isomer, salicylic acid, was conjugated with
glycine. With the m- and p-isomers, however, only a small part was conjugated with glycine, whereas
the major part appeared as the sulfuric acid conjugate. It was the first example of compounds able to use
two alternative conjugation pathways. This surprising finding brought up the idea of a possible
competition of the two conjugating agents for these substrates and, furthermore, that the formation of a
conjugate molecule would inhibit its reaction with the alternate conjugating agent. Experimentally,
Baumann and Herter then tried to increase one of the conjugating agent pools by administering either
glycine or sulfate. Contrary to their expectations, the excretion of either conjugate was not increased and
the attempt at shifting the reaction equilibrium within the organism had failed. Still, the approach was
remarkable.
A quantitative study on the conjugation of phenol was carried out by Schaffer [97] in Nencki's
laboratory in 1878. He administered phenol to dogs, collected their urine for several days, and determined the phenylsulfuric acid excreted. In all animals he thereby recovered only two-thirds of the dose, a
result soon confirmed by Tauber [98] and Auerbach [99]. Like Schultzen and Naunyn and Munk,
Schaffer thus was once more confronted with the question about the fate of the unrecovered compound.
The oxidation of benzene to phenol might serve as an indication that phenol underwent further
hydroxylation to an unknown product which presumably would also be excreted as a sulfuric acid
conjugate. His theory for an
24
CONTI AND BICKEL
experimental approach was the following. When a dog was given a dose of phenol, the excess sulfuric
acid in the urine should correspond with the excreted phenol moiety. Any further excess would then
indicate that in addition to phenol another substance had been formed and excreted as sulfate conjugate.
Such a further excess was indeed detected by Schaffer, thus suggesting the formation of an additional
sulfuric acid conjugate. This result was examined in the light of two observations made by Salkowski
who found that after intestinal ligation the urine contained sulfuric acid in excess of indican and phenol
[100], and furthermore, that phenol-poisoned rabbits excrete oxalic acid [101]. However, Schaffer was
not able to detect an increase in oxalic acid in the urine of phenol-treated dogs and thus dismissed the hypothesis of oxalic acid as a phenol metabolite. The problem was followed by Baumann and Preusse [102]
who, in analogy to the hydroxylation of benzene to phenol, postulated a hydroxylation of phenol to
catechol and hydroquinone. The sulfate conjugates of these oxidation products were regarded as
Schaffer's missing phenol metabolites. Baumann's and Preusse's hypothesis was soon proved right by
Nencki and Giacosa [103] in 1880. A dog was given a large dose of benzene per os and by simultaneous
cutaneous administration. Catechol could indeed be detected in the dog's urine. Studies in man again
yielded catechol and, in addition, trace amounts of hydroquinone. Thus Schultzen and Naunyn's
hypothesis of a further oxidation of phenol eventually proved to be correct. Nencki and Giacosa,
however, were aware that, starting from benzene, a considerable proportion still escaped detection. Their
explanation for this fact was incomplete intestinal absorption and excretion of the unchanged compound
by the lungs.
At that point benzene was the first foreign compound that had been shown to form as many as half a
dozen metabolites by stepwise metabolism along oxidative and conjugative pathways:
OSO3 H
OH
OH
OH
Catechol
Hydroquinone
OH
HO
OH
OSO3H HO
OSO3H
HISTORY OF DRUG METABOLISM
25
V. REDUCTION
The first reduction of a foreign compound was discovered 21 years after the first oxidation. It was
a quick discovery by one man who, in 1863, had knowledge of the metabolism of benzoic acid and who
had improved analytical and physiological techniques at his disposal. Speculating on chemical
similarities between quinic acid and benzoic acid, Lautemann [104] succeeded in reducing the former to
the latter by using hydrogen iodide. Once more, an analogy between chemical reagents and the organism
as a chemical agent was considered:
The fact that quinic acid was so easily reduced to benzoic acid in the test tube suggested
investigating whether the former compound was also reduced in the animal body and the resulting
benzoic acid excreted as hippuric acid. [104]
The author and two volunteers ingested 8 g of calcium quinate and collected their urines which were
subsequently analyzed. All three specimens did contain hippuric acid. This finding was soon confirmed
by Meissner and Shepard [36] in humans, rabbits, and goats; however, in contrast to Mattschersky [105],
not in dogs. At that time both the empirical formula for quinic acid, C6H7(OH)4(CO2H), and its six membered ring structure were known.
OH
HO
OH
HO
COOH
2H
-4H2O
COOH
COGly
H
Quinic acid
Benzoic acid
Hippuric acid
The studies on quinic acid metabolism were carried on in 1879 by Stadelmann [106]. His goal was
to quantify the metabolic reaction by means of hippuric acid determination according to Bunge and
Schmiedeberg and, furthermore, to clarify the species differences mentioned above. In all cases he
carefully determined hippuric acid excretion in his animals before and after quinic acid administration. In
agreement with Meissner and Shepard [36], dogs given 2 to 3 g of quinic acid showed no increase in
hippuric acid excretion. In contrast, rabbits, selected as a herbivorous species, in all cases excreted
excess hippuric acid. Two important observations were emphasized by Stadelmann:
The excess hippuric acid was far from representing
26
CONTI AND BICKEL
the quinic acid doses ingested, and its appearance in the urine occurred 24 to 48 hr after ingestion only.
The first observation was confirmed much later by Forster [107] and Hupfer [108] who, after ingesting a
total of up to 100 g of quinic acid, recovered some 20% of the dose as urinary hippuric acid. The lack of
quinic acid reduction in the dog was confirmed by Schmidt [109] who, on the other hand, found an
increase in sulfuric acid after urinary hydrolysis, indicating an unknown sulfuric acid conjugate.
Stadelmann then investigated the localization of quinic acid reduction much as Schmiedeberg,
whose disciple he was, had done in the case of glycine conjugation. A reduction in the blood was unlikely in view of the remarkable time lapse from ingestion of quinic acid to excretion of hippuric acid.
Still, the substrate was injected into the jugular vein of rabbits. As expected, no increase in hippuric acid
occurred. Reduction in the gastrointestinal tract was therefore more likely. The question here was in
which part the reaction might occur, and whether or not under the influence of a secretion product. The
first experiments were aimed at the stomach as a potential reduction site. However, pylorus ligation in
rabbits did not result in increased hippuric acid after quinic acid administration; neither did incubation
with pancreas homogenate for 18 hr at 370C. At this point Stadelmann gave up further experimentation
and offered a hypothesis which later proved right:
The site of metabolism of quinic acid is in all likeliness the intestine, although we have not
succeeded in furnishing the appropriate experimental evidence.... The retarded excretion suggests
that reduction of quinic acid occurs slowly, presumably in the distal part of the intestine...
.Whether the transformation in the lower intestine is part of the putrefactive reduction processes,
or whether it is due to the action of glandular secretions, cannot be decided yet, although the
former alternative seems to be more likely.[106]
Stadelmam, too, was harassed by the problem of low metabolite recovery. Even though he was working
with a sensitive method for the determination of hippuric acid, the urine of his rabbits yielded only a
fraction of the administered dose. Since, however, there was no method for the detection of quinic acid at
that time, he speculated that the major part of a dose was excreted as unchanged quinic acid.
Quinone was another foreign compound to be reduced in the hody. The first experiments were
carried out by Woehler and Frerichs [29]
HISTORY OF DRUG METABOLISM
27
in 1848. These authors were not able to detect quinone in the urine of dogs dosed with 0.5 to 1.0 g of the
compound which, in an era not used to drug metabolic reactions, was most surprising to them. Only in
1892 was this investigation resumed by Schulz [110] who found that urine after ingestion of quinone had
a dark brown-green color and that after hoiling with hydrochloric acid a compound was obtained which
reduced silver nitrate and was likely to be hydroquinone. Indeed, S. Cohn [111] succeeded in isolating
crystalline hydroquinone which had been excreted as its sulfuric acid conjugate:
O
HO
HO
O
Quinone
OH
OSO3 H
Hydroquinone
Hydroquinone sulfate
The last two decades of the 19th century also led to the discovery of nitro reduction. The first
example was indicated by Hoppe-Seyler [112] who found that o-nitrophenylpropiolic acid was
metabolized and excreted as indoxylsulfuric acid. This was interpreted as an initial reduction followed by
decarboxylation to indoxyl and sulfuric acid conjugation. The last reaction had already been described by
Baumann and Herter [92].
OH
OH
CH 2COOH
NO2
Indoxylic acid
COOH
N
H
o-Nitrophenylpropiolic acid
OH
N
H
Indoxyl
OSO3 H
N
H
Indoxyl sulfate
Another instance of reductive metabolism of nitro groups was the metabolic transformation of picric acid
(2,4, 6-trinitrophenol) to picramic acid (2-amino -4, 6-dimtrophenol). This reaction was postulated by
Rymsza [113] and established by Karplus [114].
The nitro reductions of nitrobenzaldehydes were of particular interest. Sieber and Smiruow [115]
reported that in dogs the three isomers of nitrobenzaldehyde were oxidized to the corresponding nitrobenzoic acid isomers which then usually were conjugated to nitrohippuric acids. R. Cohn [116], who
was also dealing with the
28
CONTI AND BICKEL
metaholism of nitrobenzaldehydes, observed that rabbits given the m- and p-isomers of that compound
excreted an unknown product in addition to the nitrobenzoic and nitrohippuric acid isomers. Cohn succeeded in identifying the new metabolite as acetylaminobenzoic acid. Whereas he first assumed the
reduction to take place in the intestine, he later also obtained the metabolite in rabbit urine after
subcutaneous administration of nitrobenzaldehyde and thus concluded that this nitro reduction was a
truly endogenous metabolic reaction [117]. Cohn was the first to demonstrate that two functional groups
in a single molecule could be metabolically attacked so that such drugs undergo two separate pathways:
O2 N
O2 N
COOH O2 N
H2 N
COOH
COGly
CHO
CH 3CONH
COOH
Among the highlights of early pharmacology and drug metabolism is the saga of chloral hydrate.
This compound was discovered in 1831 by Liebig, but only in 1861 was it studied with respect to its
pharmacological properties by Buchheim. Based on the formation of chloroform and formic acid upon
heating chloral hydrate with potassium hydroxyde, Buchheim thought he had found a drug able to
release acid in the body and thus become useful in counteracting alkalosis. Animal and selfexperimentation with the drug then disclosed the strong hypnotic action of chloral hydrate, which
Buchheim ascribed to the metabolic formation of chloroform. Unfortunately, he did not publish these
results which therefore remained in obscurity until Kobert [118] and others were able to quote from his
posthumous works. For this reason Liebreich [119] who, 8 years after Buchheim, worked and published
on chloral hydrate, is given credit for discovering its hypnotic action. Indeed, his discovery was made
without knowledge of Buchheim's work and was the starting point for the introduction of this new
synthetic hypnotic. Liebreich, too, postulated that the drug's hypnotic action is due to chloroform which
is formed by alkaline constituents of the blood. Loebisch in 1888 [120] praised Liebreich not only for the
introduction of a most successful drug, but also for being the inaugurator of modern experimental
pharmacology which synthesizes drugs based on chemical and biochemical knowledge, tests their ac-
HISTORY OF DRUG METABOLISM
29
tions in animals, and introduces them into therapy with a circumscribed indication. This statement, to be
sure, characterized new trends in pharmacology rather than Liebreich in particular, who had introduced
chloral hydrate and propagated the chloroform theory, but was unable to prove it. Even after giving high
doses of chloral hydrate, the urine of his animals was devoid of chloroform. The only positive statement
was his impression that the animals' expiration might contain chloroform.
In the following years a host of authors tried to obtain evidence for or against the chloroform
theory. Personne [121] reported detecting chloroform and formic acid in the blood of chloralized animals, although most French investigators denied Liebreich' s theory. Others, like Hammarsten [122] and
Rajewsky [123], were unable to detect chloroform in blood or expired air. Kulz [124] reported having
found chloroform, and Bouchut [125] chloral hydrate in urine. Tomaszewicz [126] added to the
confusion by reporting the presence of chloral hydrate and the absence of chloroform.
This was the state of affairs in 1875 when the classical paper of von Mering (Fig. 3) and Musculus
[127] appeared:
There is hardly a drug which in recent years was consumed in such large amounts as chloral
hydrate, and on whose mechanism of action so much controversy exists.... These contradictory
reports above all incited us to investigate the fate of chloral hydrate in the organism. [127]
Von Mering and Musculus found no chloroform in the expired air of chloralized animals. In the urine of
humans who had ingested 5 to 6 g of the drug daily for some time, small amounts of unchanged drug, but
no chloroform or formic acid was detected. This was not the end but rather the beginning of their
investigation which next disclosed that the above urine was levorotatory without containing albumin.
Tedious work eventually resulted in the isolation of the levorotatory compound which was an acid called
urochloralic acid by von Mering and Musculus. It was believed to originate from the reaction of an
unknown metaholite of chloral hydrate with an endogenous compound.
The same compound was isolated two years later by Falck [128] who also believed it to be a
metabolite of chloral hydrate and who predicted that the discovery of urochloralic acid would mark the
end of Liebreichs's chloroform theory. After several investigators had not succeeded in preparing
urochloralic acid in amounts sufficient for analytical work, Kulz [129] by 1880 made another most
remarkable contribution. By boiling urochloralic acid from the urine of chloralized dogs with
hydrochloric acid, he obtained a chlorinated compound
30
CONTI AND BICKEL
FIG. 3. Josef von Mering, 1849-1908. Strassburg, Halle.
and a dextrorotatory, strongly reducing acid. The latter proved to correspond to glucuronic acid which
had just been discovered by Schmiedeberg and Meyer [130] By administering single doses of 20 to 25 g
of chioral hydrate to dogs, Kulz [131 succeeded in isolating enough urochioralic acid to perform
elementary analysis. The empirical formula, C8H13C1307, even though obtained with two different
methods, unfortunately was not correct and created some confusion. Convinced of the correctness of
both the formulas of urochioralic and glucuronic acid, Kulz postulated the unconjugated metabolite of
chloral hydrate to be C2H5C130. He adhered to that formula, although he was aware that GarzarolliThurnlackh [132] had obtained
HISTORY OF DRUG METABOLISM
31
trichioroethanol (C2H3C130), a likely candidate for the unconjugated chioral hydrate metabolite, by
reducing the drug with ethyl zinc.
Von Mering and Musculus [127], too, had indicated a wrong formula for urochloralic acid in their work
of 1875; however, they had emphasized the formula to be preliminary, since the purity of their
preparation was not satisfactory. In 1882, the same year Kulz published his incorrect findings, von
Mering [133, 134] had reached his goal of correctly identifying urochloralic acid and its aglycone resulting from acid hydrolysis:
C8H11C1307
Urochloralic acid
+
H20
C2H3C130 +
T richloroethanol
C6H1007
Glucuronic acid
Von Mering found the properties of his aglycone to fully correspond to trichloroethanol as obtained in
the test tube by Garzarolli-Thurnlackh, and thus urochloralic acid to be trichloroethyl glucuronide. He
particularly emphasized that trichloroethanol was the primary metabolite of chloral hydrate:
With the glucuronides known so far, the corresponding aglycones originated in the organism
by oxidation of the ingested compound. In our case, however, the aglycone (trichloroethyl
alcohol) resulted from reduction. This is why urochloralic acid deserves particular interest.
[134]
Thus, 13 years after the introduction of chloral hydrate as a hypnotic and after Liebreich's chloroform
theory, the drug had eventually been shown to undergo metaholic reduction and subsequent glucuronic
acid conjugation. A final contribution was made by Kulz [135] in 1884 by demonstrating the formation
of urochloralic acid from ingested trichloroethanol in rabbits. This unequlvocally proved von Mering's
findings.
VI, GLUCURONIC ACID CONJUGATION
It has been mentioned in the previous section that in 1875 von Mermg and Musculus [127] in the course
of their work on the fate of chloral hydrate isolated urochloralic acid, an apparently conjugated
metabolite, which subsequently was characterized as a glucuronide by Kulz [129]. These discoveries,
however, were only the final stages of the process, which over three decades gradually disclosed the
mechanism of glucuronic acid conjngation.
32
CONTI AND BICKEL
This development started with Indian yellow, a dye obtained from the urine of cows fed on mango
leaves. Its main constituent, euxanthic acid, was suspected by Erdmann [136] in 1846 to be a conjugated
compound, since it formed a new acid upon hydrolysis. Schmid [137] knew that Indian yellow was
produced from the urine of camels fed on the frnits of Mangostana mangifer. In 1855 he therefore
analyzed both euxanthic acid and what he called mangostin, a yellow pigment from the mango fruit.
Obtaining similar empirical formulas for mangostin and euxanthic acid, Schmid speculated that the latter
compound might be formed in the camel from the former:
It is readily conceivable that mangostin is transformed to euxanthic acid by the vital
process.... If euxanthic acid is treated with concentrated sulfuric acid and the resulting solution
poured into water, then, as is known, euxanthone precipitates. The filtrate then easily reduces
copper oxide dissolved in potassium hydroxide, whereas neither euxanthic acid nor euxanthone
have reducing properties.
[137]
Thus the filtrate was shown to contain a substance that had been part of euxanthic acid, thereby
demonstrating that this compound was a conjugate. In addition, a first chemical property of the new
conjugating agent had been discovered. Although Schmid did not mention whether he was familiar with
glycine conjugation, this is strongly suggested by his experimental procedure.
After Schmid's paper had appeared, several other investigators dealt with euxanthone and
proposed a number of empirical formulas. Real progress was not achieved before Baeyer [138] in 1870
gave the first correct formula for euxanthone and verified Schmid' s findings, including the copper oxidereducing conjugating agent. Based on accurate analyses, he then was able to indicate the correct
relationship between euxanthone and euxanthic acid, which in Baeyer’s empirical formulation and in
modern structural formulas reads:
C19H16O10 + H2O = C6H10O7 + C13H8O4
COOH
O
OH
O
COOH
OH
O
OH
O
HO
OH
Euxanthic acid
(Indian yellow)
O
O
OH
+
HO
OH
Glucuronic acid
OH
HO
O
Euxanthone
HISTORY OF DRUG METABOLISM
33
Baeyer then tried to characterize the unknown conjugating agent, C6H10 07, obtained by hydrolysis of
euxanthic acid. In addition to its reducing power he was aware of Laurent's observation that upon
heating, euxanthic acid decomposes into euxanthone, carbon dioxide, and water. Both the reduction and
the type of decomposition were reminiscent of sugars. Furthermore, the empirical formula of the
conjugating agent almost corresponded to that of saccharic acid, C6H1008. Based on these pieces of
evidence, Baeyer proposed that the conjugating agent was kind of a saccharic acid and thus euxanthic
acid had to be considered as a glycoside of saccharic acid with euxanthone.
When von Mering and Musculus [127] in 1875 isolated urochloralic acid as a metabolite of chloral
hydrate in human urine, they were mainly interested in the chlorine-containing moiety of the molecule.
Yet they also contributed to the characterization of the conjugating agent by comparing the conjugate,
urochloralic acid, with the parent drug. Thus, in agreement with Baeyer and Schmid, urochloralic acid
was found to be considerably acidic and able to reduce boiling alkaline copper solution and silver and
bismuth oxides. Furthermore, the conjugate possessed levorotatory optical activity and, upon boiling
with potassium hydroxide, turned brown and developed a smell of caramel. Von Mering and Musculus
concluded from these findings that urochloralic acid could be split into a chloral hydrate derivative
(which later turned out to be trichloroethanol) and an endogenous, sugarlike compound. Being familiar
with glycine conjugation, they were convinced that they were dealing with an unknown type of
conjugation. They also mentioned having found other substances forming reducing and levorotatory
urinary metabolites such as morphine and crotonchloral hydrate, thereby suggesting that all these drugs
undergo a common conjugation reaction.
When dogs or rabbits were poisoned with nitrobenzene, Ewald [139] found that their urines contained a
reducing compound which he believed to be sugar. He therefore suggested the toxicity of nitrobenzene to
be due to diabetogenic action. This, however, was disproved by von Mering [140] who demonstrated that
the urine of nitrobenzene-treated rabbits contained a levorotatory compound rather than dextrorotatory
glucose. Using the same model, Jaffe [141] in 1878 was not able to find optical activity and reducing
power in all his experiments. He questioned whether the urinary metabolite was a derivative of
nitrobenzene and offered the hypothesis that the metabolite originated from nitrotoluene, a possible
contaminant of commercial nitrobenzene. For a number of years Jaffe had been investigating the
toxicology and metabolism of the three isomers of nitrotoluene. He was particularly curious to know
whether all isomers were
34
CONTI AND BICKEL
metabolized like toluene, or else, whether their different chemical properiies were reflected in different
metabolic pathways. In a first paper, Jaffe [63] reported that besides p-nitrotoluene, the urine contained
both p-nitrobenzoic acid and its glycine conjugate, p-nitrohippuric acid. Four years later [141] he stated
that, in contrast to the p-isomer, o-nitrotoluene in the dog resulted in ouly 10% of o-nitrohippuric acid,
the major part being a more complex conjugate. Due to certain similarities with urochloralic acid, Jaffe
called his conjugate uronitrotoluic acid. Starting out from a correct empirical formula of the conjugate,
and based on the assumed analogy with the p-isomer as well as a sugarlike conjugating agent, Jaffe first
proposed Reaction I. However, neither nitrobenzoic acid nor a hexose could be identified after
hydrolysis. Rather, the aglycone was found to be nitrobenzyl alcohol. This then led to the correct
Reaction II:
I (false):
II
C13H15N09 + H20
Uronitrotoluic
acid
(correct): C13H15N09 + H20
Uronitrotoluic
acid
C7H5N04 + C6H1206
Nitrobenzoic Sugar
acid
C7H7N03 + C6H1007
Nitrobenzyl "Sugar"
alcohol
Jaffe now paid particular attention to the conjugate, C6H1007, which he cautiously called "hypothetical
acid." Like some of his predecessors, he characterized it as a reducing dextrorotatory compound which,
upon heating, developed a caramel-like smell. These were obviously properties of carbohydrates.
However, sugar could be ruled out due to the empirical formula as well as to the observation that the
compound did not undergo fermentation by yeast. Jaffe concluded that the conjugating agent was an
oxidation product of sugar, probably formed by transformation of an -CH2OH into a -COOH group. By
these correct conclusions Jaffe indeed came very close to glucuronic acid.
Camphor was still another drug forming a metabolite which was composed of a camphorlike and a
sugarlike moiety. This observation was made by Wiedemann [142] who boiled the urine of camphor treated dogs with dilute acids. He speculated that the metabolite might be a nitrogen-containing
glycosidic acid. This study was resumed by Schmiedeberg (Fig. 4) and Meyer [130] who, in 1879, succeeded in solving the problem of the sugarlike conjugating agent. They reproduced the formation of the
conjugated camphor metabolite in dogs as well as the hydrolysis of the conjugate. With relative ease
HISTORY OF DRUG METABOLISM
35
FIG. 4. Oswald Schmiedeberg, 1838-1921. Dorpat, Strassburg.
they were able to extract and purify the camphorlike aglycone which they characterized as C10H16O2, a
dextrorotatory secondary alcohol; i.e., an oxidation product of camphor with an additional alcohol group.
This primary metabolite was therefore named campherol.
More difficulties were encountered by Schmiedeberg and Meyer during the identification of the
conjugating agent. They were the first to eventually succeed in isolating a small amount of the pure
conjugating acid. This then allowed detailed investigation of the compound which led to the following
conclusions:
There can be no doubt that this acid is a direct derivative of glucose. This is suggested by
the formula, the reduction of alkaline copper solution, the dextrorotation of the plane of
polarization, and the results of oxidation experiments which exclude the participation of an
aromatic nucleus in its formation. Based on the composition of this acid, its structure is
intermediate between gluconic acid and saccharic acid....
36
CONTI AND BICKEL
According to Fittig's views on the structure of glucose, the acid must be given the formula
(CH.OH)4
CHO
COOH...
Arbitrarily we call the compound glucuronic acid. [130]
Thus Schmiedeberg and Meyer established the metabolism of camphor as a primary oxidation to
campherol, followed by glucuronic acid conjugation which resulted in the formation of
camphoglucuronic acid.
Finally, Schmiedeberg and Meyer also speculated on the origin of glucuronic acid in the organism. They
strongly suspected that their discovery allowed insight into the mechanism of glucose oxidation. Since
acids had been postulated as early intermediary metabolites of glucose, glucuronic acid was a likely
metabolite which, by conjugating with campherol, had escaped furiher breakdown and combustion. Thus
experiments with foreign compounds like camphor could also be useful in investigating processes of
intermediary metabolism.
VII. ACETIC ACID CONJUGATION
The finding of acetylaminobenzoic acid as a metabolite of nitrobenzaldehyde [116] has been reported in
the section on metabolic reductions. The actual discovery of acetic acid conjugation took place six years
earlier, in 1887, when Jaffe and R. Cohn [143] described what still is known as a most unusual metabolic
reaction involving acetic acid. Based on chemical similarities of benzene and furan, the authors
investigated the fate of furfural in dogs and rabbits. The urine of the animals contained α-furoic acid
(called pyromucic acid, C5H403), furoylglycine (pyromucuric acid), and a third acid metabolite. The
latter could be split by hydrolysis into glycine and a nitrogen-free acid with the empirical formula C7
H603. A compound of the same empirical formula had been obtained ten years earlier by Baeyer [144]
after boiling furfural and sodium acetate for 8 hr. He correctly called the product furfuracrylic acid. Jaffe
and Cohn carefully compared their nitrogen-free acid with Baeyer's furfuracrylic acid and found identity
of all chemical properties tested. This was further confirmed by administering the acid to rabbits which
then excreted the glycine conjugate. Thus furfural in the organism must have undergone a synthetic
reaction with acetic acid, thereby forming furfuracrylic acid, which subsequently was conjugated with
glycine. Jaffe and Cohn expressed their surprise ahout this unheard of
HISTORY OF DRUG METABOLISM
37
metaholic reaction which was thought to shed new light on biosynthetic processes at large. Their results
on the metabolism of furfural can be formulated as follows:
COOH
O
COGly
O
Furoylglycine
CHO
O
Furfural
O
C CHCOOH
H
O
C CHCOGly
H
Furfuracrylic acid
Obviously, this discovery justified further investigation with additional aldehydes. R. Cohn [116] used
α-thiophene aldehyde, the thio analog of furfural, as another substrate to demonstrate the biosynthetic
process with acetic acid. The urine of rabbits and dogs treated with the substrate contained thiophenuric
acid, the glycine conjugate of α-thiophenic acid. However, it did not contain thienylacrylic acid, the expected analog of furfuracrylic acid. On the other hand, injection of synthetic thienylacrylic acid resulted
in the excretion of
α-thiophenic acid without unchanged compound in the urine of a rabbit, thus indicating the metabolic
cleavage rather than the biosynthesis of thienylacrylic acid:
COGly
S
Thiophenuric acid
CHO
S
Thiophene
aldehyde
S
C CHCOOH
H
S
C CHCOGly
H
CH3COOH
Thiophenylacrylic acid
R. Cohn was so convinced of the formation of thienylacrylic acid that he explained its lack in the urine
by its rapid breakdown into α-thiophenic acid. Actually, by discovering the metabolic formation of furfuracrylic acid from furfural, R. Cohn had come across an atypical biosynthesis with acetic acid. This
made him believe acetic acid conjugation to be a reaction between an aldehyde and the methyl group of
38
CONTI AND BICKEL
acetic acid. His stubborness in testing suitable aldehydes eventually led him to discover the typical acetic
acid conjugation of amines:
Atypical:
R-CHO + CH3-COOH
R-CH=CH-COOH
Typical:
R-NH2 + HOOC-CH3
R-NH-CO-CH3
Nitrobenzaldehydes became the crucial substrates for R. Cohn's discovery of the typical acetic acid
conjugation. Again, the first experiments were disappointing; using o-nitrobenzaldehyde, R. Cohn, like
Sieber and Smirnow [115], found o-nitrobenzoic acid in the urine, but no conjugate whatsoever. A dog
given m-nitrobenzaldehyde excreted m-nitrohippuric acid without acetic acid conjugate. However, using
rabbits instead of dogs, R. Cohn obtained an unknown acid with an empirical formula C9H9N03.
Hydrolytic cleavage of this metabolite yielded m-aminobenzoic acid and acetic acid. The underlying
metabolic process was correctly interpreted by R. Cohn:
Concerning the formation of acetylaminobenzoic acid from nitrobenzaldehyde in the animal
body, one must assume that nitrobenzoic acid, which is primarily formed, is reduced to
aminobenzoic acid in the long intestine of the rabbit, and that the latter compound is absorbed and
finally in some organ meets acetic acid:
C6H4
NH2 + CH3.COOH
COOH
C6H4
NH.CO.CH3 + H20
COOH
...I have demonstrated in the above communication that syntheses with acetic acid play an imporiant
role in the animal body. Acetic acid is thus frequently involved in metabolic reactions, even
though it usually escapes detection due to its rapid oxidation. [116]
As has been mentioned in the section entitled "Reduction," R. Cohn [117] later found that the nitro
reduction took place in organs rather than in the intestinal lumen. Thus R. Cohn's work was not only a
remarkable contribution to the history of drug metabolism but, as demonstrated by the end of the above
quotation, it also introduced acetate, one of the most important intermediary metabolites, into biochemistry.
HISTORY OF DRUG METABOLISM
39
VIII. GENERAL COMMENTS
This work is restricted to the discoveries of the major metaholic pathways within the history of drug
metabolism in the 19th century. Since the main events described in Sections II to VII are summarized
below, this section attempts to discuss and comment upon a number of individual aspects. At times, this
is deliberately done from the viewpoint of modern biochemistry.
The previous sections deal with the history of drug metabolism or xenobiochemistry during the period
1840 to 1900. What was achieved during that period of early biochemistry in our particular field? Above
all it brought about knowledge of those metabolic pathways of xenobiotics which still today are
considered the most important ones: oxidation, reduction, and conjugations with glycine, sulfuric acid,
glucuronic acid, and acetic acid. The wealth of individual studies and results published during that period
was quite remarkable, but this particular aspect is beyond the purpose of this work. Suffice it therefore to
mention Heffter's [7] monograph of 1905, which reviews the fate in the organism of over 400 organic
compounds and lists no less than 600 references. It characterizes the time and the stage of development
of biochemistry when Heffter uses the classification of his compounds according to the venerable
Beilstein Handbook, when "foreign to the hody" is used synonymously with "foreign to the urine," when
many compounds like fatty acids, amino acids, and other intermediary metaholites are treated as foreign
compounds.
Dealing with 19th century drug metabolism, one should bear in mind that important dimensions like
enzymology, chemical reaction mechanisms, pharmacokinetics, and cell biology were introduced in the
course of the 20th century. The fact that 19th century biochemistry was preenzymological is of particular
importance. This precluded a real distinction between chemistry or test tube chemistry on the one hand
and biochemistry as a chemistry of selected compounds occurring in organisms and determined by
genetically coded enzymes on the other hand. Still, within that frame, biochemistry, which started at a
near-zero level of knowledge around 1800, became a remarkable success story in the course of its first
century. The factors limiting its progress were chiefly the respective state of analytical and physiological
techniques. Many brilliantly conceived projects were doomed to failure for lack of sufficiently sensitive
analytical methods.
Who were the investigators who decided to study the metabolism of foreign compounds, and what was
their motivation? By training,
40
CONTI AND BICKEL
they were chemists as well as physiologists, pharmacologists, and clinical physicians. Accordingly, their
outlooks and approaches varied at times. Clinically relevant problems were relatively rare at that time.
The same holds for the interest in foreign compounds as drugs, since the amount and importance of
synthetic drugs was still modest and the notion of environmental contaminants was nonexistent. Thus it
was biochemistry which mainly motivated research in the field. The discovery of the biotransformation
of benzoic acid to hippuric acid was a major event since it suggested that metabolic pathways and
processes could be investigated by means of simple foreign compounds. This was a tempting alternative
to the use of the complex nutrients and body constituents, which in most instances were poorly
characterized. Many investigators using a chemical approach dealt with the organism as a chemical
reagent which allowed them to study new reactions.
Most researchers used the same type of basic methodology in order to obtain information on the
biochemical fate of a drug. The compound was administered per os to a lahoratory animal or to a
volunteer, and the urine was qualitatively analyzed for unchanged drug or its metabolite. Very rarely
were blood or tissue samples analyzed, since the available methods lacked the sensitivity for the
detection of the small amounts of drug which had been diluted in the large volume of the organism. In
most cases the doses used in an experiment were limited because of the unpleasant or toxic effects of the
compounds. A most striking feature was the habit of self-experimentation, performed by many
investigators. No less impressive are the "heroic" doses used, which often were in the multigram range.
Many critical remarks can be made with respect to the art of conducting experiments in those days. The
following by no means is meant to be a critique of the pioneers, underprivileged in terms of information
and technical facilities, but rather as a means of characterizing their time and their situation. For
example, the methods used often precluded the detection of several metabolites of a drug, and only
allowed the detection of what had been expected or suspected. Most experiments were carried out in
vivo, i.e., in an extremely complex and largely unknown system. Pharmacokinetic processes like
absorption, distribution, tissue uptake, extrarenal excretion, though closely associated with metabolism,
were frequently neglected. Also neglected were species differences; in fact, many well-conceived
experiments were carried out in unsuitable species, or else results were briskly extrapolated to other
species. Quantitative determinations, if at all considered, were hampered by poor accuracy and
sensitivity of analytical methods. Decisive control experiments were omitted at times; e.g., not all
hippuric acid determinations were checked against hippuric
HISTORY OF DRUG METABOLISM
41
acid produced from endogenous aromatic compounds. Recoveries often comprised a minor fraction of
the dose, which then led to the conclusion that the remaining drug had been degraded to carbon dioxide
and water. Certain researchers were very quick in making a grandiose biological principle out of one or a
few experimental results. Finally, it took biochemists the whole 19th century to quit thinking in terms of
organic chemical test tube reactions.
However, a number of works were produced in drug metabolism, which in terms of conception, quality,
and impact were far ahead of their time and must be regarded as pacemakers of modern biochemistry.
These achievements are highly respectable in view of the "preenzymological" era and the primitive
equipment of the laboratories. Many of the important discoveries made in xenobiochemistry were aiming
at or have solved complex and difficult problems. It has been mentioned that the conjugating agent
glucuronic acid was gradually unraveled by a number of investigators over the period 1846 to 1879, and
that its basic structure was established long before the complete structure and the metabolism of glucose
was known. In addition to a host of simple metabolic reactions, multiple pathways of xenobiotics were
also studied or even provoked with suitable substrates. Of particular importance was the successful
elucidation of reaction sequences like oxidation conjugation, which has become the backbone of
xenobiochemistry. Within series of aromatic or other compounds, scores of individual substances were
studied so as to allow the establishment of rules on their metabolic fate. The popularity of aromatic
compounds in the field of drug metabolism was due to their easy recognizability and to the fact that,
except for aromatic amino acids, they hardly occur in the organism and are not metabolized to the extent
of rupturing the benzene ring. Therefore, the benzene ring was used as a metabolically stable label and
experiments were conceived which are reminiscent of those with radioactive labels performed in our
time. Experimental failures often were transformed into provocative and stimulating hypotheses. For
example, hydrolyses like digestive processes, but not biosyntheses like hippuric acid formation, could
easily be stimulated in vitro. This dilemma provided a strong stimulus to develop in vitro systems and
thereby to study the conditions prevailing in vivo. In the case of the above biosyntheses, this may even
have been a last opportunity to reflect on the "vis vitalis." Another type of experiment which did not
yield the expected answer was the attempt at influencing what was supposed to be a competition of
conjugation reactions. But again, this stimulated the conception of limited pools of endogenous
conjugating agents, like glucuronate and sulfate. The use of knowledge in drug metabolism to improve
pharmacodynamic properties of
42
CONTI AND BICKEL
drugs anticipated certain aspects of what in the second half of the 20th century be came known as
molecular pharmacology. Furthermore, certain investigations of the localization of xenobiochemical
processes must be called spectacular at a time which hardly had abandoned the notion of the blood as the
site of general metabolism. Some of these localization studies were performed with new and bold
techniques whereby biochemical processes were explored under well-defined conditions of physiological
alterations. Among these new techniques were the bile fistula which eliminated a secretion, nephrectomy
which eliminated an organ, the isolated perfused organ eliminating undesired influences of the whole
organism, the ligated liver, and the portocaval shunt which was then called Eck's fistula. Among the
achievements of the time there was even the use of "tool" drugs which were given to alter physiological
parameters for the study of a test drug.
There can be no doubt that studies on the metabolism of foreign compounds, which in the 19th century
was a modest speciality, has made major contributions to general biochemistry. For example, the
formation of hippuric acid from glycine and benzoic acid was the first example of a biosynthetic process
and thus introduced a concept of fundamental importance. Similar to this conjugation process, the oxidation of xenobiotics like benzene could only be simulated outside the organism by using chemical tricks,
such as ozone as an oxidant. Studies along that line strongly stimulated interest in the general process of
biological oxidation, and it was precisely the use of foreign compounds which provided a promising tool
for further investigation. Thus there is a direct line leading from benzene oxidation to the clinically used
"benzene test" and the ozone theory to the concept of active oxygen and eventually to the hypothesis of
labile, autoxidizable cell proteins as catalysts for the oxidation of endogenous and foreign compounds.
These efforts have also accelerated the rise of the concept of tissues rather than blood as metabolic sites.
The study of metabolic reductions then demonstrated that, in addition to tissues, the intestinal lumen was
still another site of xenobiochemical reactions. The conjugation reactions with glycine, glucuronic acid,
and acetic acid were all discovered with foreign compounds. It was only later that these processes were
found to be part of the intermediary metabolism of endogenous substrates. Many of the discoveries made
in the field of drug metabolism in those days had their consequences in general biochemistry or
pharmacology at a much later stage. As an example, Nencki's and Salkowski's studies on the sidechain
oxidation of aromatic compounds resulted in Knoop's discovery, forty years later, of the principle of
beta-oxidation of fatty acids. Similarly, further studies on acetylation of drugs led to the discovery of
coenzyme A in 1951.
HISTORY OF DRUG METABOLISM
43
Finally, 19th century investigators recognized and dealt with several problems of drug metabolism which
still are studied today and still pose considerable difficulties despite the immense increase in knowledge
and research technology.
IX SUMMARY-TABLES OF KEY DATES
A.
1773
1801
1824
1829
1830
1835
1841
1842
1844
1845
1857
1863/66
1875
1876
1877
1879
Glycine Conjugation*
Rouelle: Detection of "B" in urine of cows.
Scheele, Proust: "B" in human urine. "B" not identical with B?
Woehler: B in dogs excreted as unchanged "B."
Liebig: "B" from equine urine contains nitrogen. New compound called hippuric acid.
Woehler: Hypothesis of biotransformation of B to H.
Lehmann: H in urine of a diabetic patient.
Ure: Biotransformation experiment in humans shows B to be excreted as H.
Keller: Confirmation of B
H in humans.
Liebig: H also a normal constituent of human urine.
Dessaignes: Identification as benzoylglycine and synthesis of H.
Kuhne, Hallwachs: H formation in vivo from B and glycocholic acid. Site: hepatic vascular
system?
Meissner, Shepard: H formation in kidneys?
Spengel: Exogenous Gly used for H formation in vivo.
Bunge, Schmiedeberg: H formation localized in kidneys.
Hoffmann: CO and quinine inhibit H formation in isolated perfused liver.
Koch: H formation in kidney homogenate.
B. Oxidation
1842
1842
1848
Ure: Cinnamic acid excreted as hippuric acid.
Erdmann, Marchand: Other hippuric acid forming substrates. Primary oxidation?
Woehler, Frerichs: Benzaldehyde excreted as hippuric acid. Oxidation-glycine conjugation
sequence.
*B = benzoic acid; H = hippuric acid, "B" hippuric acid believed to be benzoic acid.
44
1851
1867
1870-90
1876
1880ff
CONTI AND BICKEL
Staedeler: Hypothesis of benzene oxidation to phenol.
Schultzen, Naunyn, Graebe: Fate of aromatic compounds.
Benzene ~ phenol, toluene ~ hippuric acid, xylene ~ toluric
acid, etc. Aromatic hydroxylations and glycine conjugations.
Nencki: Elahoration of aromatic xenobiochemistry. Rules.
Metabolically stable benzene ring as marker.
Munk: Benzene oxidation as a tool for studies in biological
oxidation.
Nencki, Sieber: Benzene test for metabolic oxidation capacity. Theories of biological oxidation (active oxygen,
autoxidizable cell proteins).
C. Sulfuric Acid Conjugation
1867
1876
1876
1877
1878/80
1881
1882
Buliginsky: Appearance of phenol in hydrolyzed urine.
Munk: "Phenol-forming substance" as benzene metabolite. New
type of conjugation?
Baumam: "Indigo-forming substance" on hydrolysis yields
sulfuric acid. Ether sulfates as conjugates of xenobiotics.
Baumann, Herter: Sulfuric acid conjugations of phenols.
Hydroxybenzoic acids as substrates for conjugation with
glycine and/or sulfuric acid.
Schaffer; Baumann, Preusse; Nencki, Giacosa: Phenol ~
hydroxyphenols ~ sulfates.
Schaniedeberg: Hypothesis of conjugation-oxidation sequence.
Nencki: Proof of oxidation-conjugation sequence.
D. Reduction
1863
1875/82
1879
1892/93
Lautemann: Quinic acid excreted as hippuric acid. Primary
reduction.
Von Mering, Musculus; Kulz: Chloral hydrate reduced to
trichloroethanol and conjugated to "urochloralic acid."
Stadelmann: Quinic acid reduction in intestinal lumen.
1882/92
Hoppe-Seyler; Karplus; R. Cohn: Reduction of
nitro to amino compounds. Localization in tissues.
Schulz, S. Cohn: Qulnone ~hydroqninone ~ sulfate.
E. Glucuronic Acid Conjugation
1846
Erdmann: Euxanthic acid (Indian yellow) is a conjugate with
an acid moiety.
HISTORY OF DRUG METABOLISM
1855
1870
1875
1878
1879
Schmid: Hydrolysis of euxanthic acid yields euxanthone and a reducing conjugant.
Baeyer: Euxanthic acid conjugating agent is C6H1007. Glycoside -like conjugate?
Von Mering, Musculus: Urochloralic acid conjugating agent is acidic, reducing, sugarlike
compound with optical activity.
Jaffe: o-Nitrobenzyl alcohol conjugated with acidic, reducing, dextrorotatory,
nonfermentable, oxidized sugar derivative.
Schmiedeberg, Meyer: Isolation and identification of the
CHO
conjugating agent for camphor: (CHOH)4
COOH
called glucuronic acid
F.
1887
1893
45
Acetic Acid Conjugation
Jaffe R. Cohn: Furfural forms glycine conjugate of furfuracrylic acid, a reaction product
with acetic acid
(R=CH-COOH)
R. Cohn: m- and p-Nitrobenzaldehyde excreted as N-acetylaminobenzoic acid. Sequence:
aldehyde oxidation, nitro reduction, acetic acid conjugation of amino group.
REFERENCES
[1]
[2]
[3]
[4]
[5]
[6]
[7]
[8]
F. Lieben, Geschichte der physiologischen Chemie, Deuticke, Leipzig, 1935.
M. Florkin, "A History of Biochemistry,"in Comprehensive Biochemistry, Vol.30 (M. Florkin
and E. H. Stotz, eds.), Elsevier, Amsterdam, 1972.
Ref. 2, Vol. 31 (1975).
R. Buchheim, Lehrbuch der Arzneimittellehre, Voss, Leipzig, 1859, p. 19.
S. Frankel, Die Arzneimittelsynthese auf Grundlage der Beziehungen zwischen chemischem
Aufbau und Wirkung, Springer, Berlin, 1901, pp.105-129. More detailed in 3rd ed., 1912, pp.
49-58, 159-206, 817-823.
A. Heffter, Ergeb. Physiol., 2, 95 (1903).
A. Heffter, Thid., 4, 184(1905).
R. T. Williams, Detoxication Mechanisms, 2nd ed., Chapman &Hall, London, 1959, pp. 13-22.
46
[9]
[10]
[11]
[12]
[13]
[14]
[15]
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]
[25]
[26]
[27]
[28]
[29]
[30]
[31]
[32]
[33]
[34]
[35]
[36]
[37]
CONTI AND BICKEL
R. L. Smith and R. T. Williams, "History of the Discovery of the Conjugation Mechanisms," in
Metabolic Conjugation and Metabolic Hydrolysis, Vol. 1 (W. H. Fishman, ed.), Academic, New
York, 1970, p. 1.
T. C. Butler, Ann. N. Y. Acad. Sci., 179, 502 (1971).
M. H. Bickel, Marceli Nencki 1847-1901, Huber, Bern 1972, pp. 33-53.
T. Thompson, A System of Chemistry, Vol.4, Edinburgh, 1802, p. 447.
W. Hallwachs, Ann. Chem. Pharm., 105, 207 (1858).
J. Berzelius, Lehrbuch der Chemie, Vol.9, 3rd ed., Dresden, 1840, p. 425.
A. F. de Fourcroy and L. N. Vauquelin, Ann. Chim. Phys., 31, 63 (1799).
L. Proust, Ibid., 36, 273 (1801).
F. Woehler, Tiedemann's Z. Physiol., 1, 142 (1824).
J. Liebig, Poggendorff's Ann. Phys. Chem., 17, 389 (1829).
0. L. Erdmann, J. Praki. Chem., 13, 422 (1838).
J A. Dumas, Ann. Chim. Phys., 57, 331 (1837).
V. Dessaignes, C. R. Acad. Sci. (Paris), 21, 1224 (1845).
V. Dessaignes, J. Pharm. (Paris), 32, 44 (1857).
A. Ure, Pharm. J. Transact., 1, 24(1841).
C. G. Lehmann, J. Praki. Chem., 6, 113 (1835).
F. Woehler, quoted by J. Berzelius, Lehrbuch der Chemie, Vol.4, Dresden, 1831, p. 376.
W. Keller, Ann. Chem. Pharm., 43, 108 (1842).
J Liebig, Ibid., 50, 161 (1844).
W. Kuhne and W. Hallwachs, Virchow's Arch. Pathol. Anat., 12, 386 (1857).
F. Wdhler and F. T. Frerichs, Ann. Chem. Pharm., 65, 335 (1848).
A. Strecker, Ibid., 65, 1, 130 (1848); 67, 1 (1848).
T. Schwann, du Bois-Reymond's Arch. Anat. Physiol., 1844,
127.
N. Blondlot, Mem. Soc. Sci. (Nancy), (1851).
A. Strecker, Ann. Chem. Pharm.,70, 149 (1849).
F. Hoppe-Seyler, J. Prakt. Chem., 89, 283 (1863).
G. Bunge and 0. Schmiedeberg, Naunyn-Schaniedeberg's Arch. Exp. Pathol. Pharmakol., 6, 233
(1876).
G. Meissner andC. U. Shepard, Untersuchungen uber das Entstehen der Hippursaure im
tierischen Organismus, Hannover, 1866.
C. Ludwig and A. Schmidt, Arb. Physiol. Anstalt Leipzig, 1868, 1.
HISTORY OF DRUG METABOLISM
[38]
[39]
[40]
[41]
[42]
[43]
[44]
[45]
[46]
[47]
[48]
[49]
[50]
[51]
[52]
[53]
[54]
[55]
[56]
[57]
[58]
[59]
[60]
[61]
[62]
[63]
[64]
[65]
[66]
[67]
[68]
[69]
[70]
47
L. Spengel, "Ueber den chemischen Vorgang bei der Bildung der Hippursaure im
Tierkdrper," Thesis, Bern, 1875.
A. Hoffmann, Naunyn-Schmiedeberg's Arch. Exp. Pathol. Pharmakol. , 7, 233 (1877).
W. Koch, Pfluger'sArch. Physiol., 20,64(1879).
C. Binz, Arch. Mikrosk. Anat., 3, 383 (1867).
C. Binz, Virchow's Arch. Pathol. Anat., 46, 129(1869).
Scharrenbroich, "Das Chinin als Antiphlogisticum," Thesis,
Bonn, 1867.
A. Ure, quoted by J. Liebig, Ann. Chem. Pharm., 44, 345 (1842).
0, L. Erdmann and R. F. Marchand, Ibid., 44, 344 (1842).
0. L. Erdmann and R. F. Marchand, J. Prakt. Chem., 26,
491 (1842).
0. L. Erdmann and R, F. Marchand, Ibid., 35, 307 (1845).
G. Staedeler, Ann. Chem. Pharm.,77, 17(1851).
A. Schlieper, Ibid., 59, 1 (1846).
0. Schultzen and B. Naunyn, du Bois-Reymond's Arch. Anat. Physiol., 1867, 349.
L. Carius, Ann. Chem. Pharm., 148, 50(1868).
M. Nencki, du Bois-Reymond's Arch. Anat. Physiol., 1870,
399.
A, W, Hofinam, Ann. Chem. Pharm., 74, 342 (1850).
C. Kraut, Thesis, Gottingen, 1854.
C. Kraut, Ann. Chem. Pharm., 98, 360 (1856).
R, Fittig and J, Konig, Ibid., 144, 277 (1867).
M. Nencki and E. Ziegler, Ber. Dtsch. Chem. Ges., 5, 749 (1872).
R, Buchheim, Arch. Physiol. Heilkunde, [N. F.]1, 122 (1857).
0. Jacobsen, Ber. Dtsch. Chem. Ges., 12, 1512 (1879).
M. Nencki, Naunyn-Schmiedeberg's Arch. Exp. Pathol. Pharmakol., 1, 420 (1873).
0. Schultzen and C. Graebe, du Bois-Reymond's Arch. Anat. Physiol., 1867, 166.
C, Bertagnini, Ann. Chem. Pharm., 78, 100 (1851).
M. Jaffe, Ber. Dtsch. Chem. Ges., 7, 248(1873).
C, Bertagnini, Ann. Chem. Pharm., 97, 248 (1856).
F. Beilstein and F. Schlun, Ibid., 133, 239 (1865).
C, Berg, "Dc nonnullarum materiarum in urinam transitudisqnisitiones," Thesis, Dorpat,
1858.
I. Munk, Pfluger's Arch. Physiol., 12, 142 (1876).
H. Landolt, Ber. Dtsch. Chem. Ges., 4, 770 (1871).
M. Nencki and N, Sieber, J. Prakt. Chem., 26, 1 (1882).
M. Nencki and N. Sieber, Ibid., 24, 498(1881).
48
[71]
[72]
[73]
[74]
[75]
[76]
[77]
[78]
[79]
[80]
[81]
[82]
[83]
[84]
[85]
[86]
[87]
[88]
[89]
[90]
[91]
[92]
[93]
[94]
[95]
[96]
[97]
[98]
[99]
[100]
[101]
[102]
[103]
[104]
CONTI AND BICKEL
M. Nencki and N. Sieber, Pfluger's Arch. Physiol., 31, 319 (1883).
0 Schultzen and L. Riess, Ann. Charite-Krankenh., (Berlin), 15, 57 (1869).
H. Quincke, Virchow's Arch. Pathol. Anat., 54, 537 (1872).
A. Schmidt, Ueber Ozon im Blute, Dorpat, 1862; Hamatologische Studien, Dorpat, 1865.
C. F. Schonbein, Sitzungsber. K. Akad. Wiss. (Munich), 1, 274 (1863).
R. Clausius, Poggendorff's Ann. Phys. Chem., 121, 250 (1864).
M, Traube, Theorie der Fermentwirknngen, Berlin, 1858.
C. Binz, Berl. Kim. Wochenschr., 9, 638 (1872).
E. Pfluger, Pfluger's Arch. Physiol., 6, 43 (1872); 10, 251 (1875).
F. Hoppe-Seyler, Z. Physiol. Chem., 1, 396 (1877).
C. Kaufmann, J, Prakt. Chem., 17, 79 (1878).
M. Nencki and P, Giacosa, Z. Physiol. Chem., 4, 339 (1880).
B. Radziszewski, Ann. Chem. Pharm., 203, 305 (1880).
A. Buliginsky, Hoppe-Seyler's Med. -Chem. Untersuchungen, 2, 234 (1867).
F. Hoppe-Seyler, Pfluger's Arch. Physiol., 5, 470 (1872).
G. Staedeler, Ann. Chem. Pharm., 144, 295 (1867).
E. Baumann, Pfluger's Arch. Physiol., 12, 63 (1876).
E. Baumann, Ibid., 12, 69 (1876).
E. Baumann, Ber. Dtsch. Chem. Ges., 9, 54 (1876).
E. Baumann, Pfluger's Arch. Physiol., 13, 285 (1876).
E. Baumann, Ber. Dtsch. Chem. Ges., 9, 1389 (1876).
E. Baumann and E. Herter, Z. Physiol. Chem., 1, 244 (1877).
B. Schuchardt, Arch. Pharm., 156, 144(1861).
Sonnenkalb, Anilin und AnilinfarbThn, Leipzig, 1864.
0. Schmiedeberg, Naunyn-Schmiedeberg's Arch. Exp. Pathol. Pharmakol. , 8, 1 (1878).
0. Schmiedeberg, Ibid., 14, 288 (1881).
F. Schaffer, J. Prakt. Chem., 18, 282 (1878).
E. Tauber, Z. Physiol. Chem., 2, 366 (1878).
A. Auerbach, Virchow's Arch. Pathol. Anat., 77, 226 (1879).
E. Salkowski, Ibid., 73, 409 (1878).
E. Salkowski, Pfl~ger's Arch. Physiol., 5, 335 (1872).
E. Baumann and C. Preusse, Z. Physiol.Chem., 3, 156 (1879).
M. Nencki and P, Giacosa, Ibid., 4, 325 (1880).
E. Lautemann, Ann. Chem. Pharm., 125, 9 (1863).
HISTORY OF DRUG METABOLISM
[105]
[106]
[107]
[108]
[109]
[110]
[111]
[112]
[113]
[114]
[115]
[116]
[117]
[118]
[119]
[120]
[121]
[122]
[123]
[124]
[125]
[126]
[127]
[128]
[129]
[130]
[131]
[132]
[133]
[134]
[135]
49
P. Mattschersky, Virchow's Arch. Pathol. Anat., 28, 538 (1863).
E. Stadelmann, Naunyn-Schmiedeberg's Arch. Exp. Pathol. Pharmakol. , 10, 317 (1879).
W, Forster, Thesis, Breslau, 1900.
F. Hupfer, Z. Physiol. Chem., 37, 302 (1903).
J Schmidt, Biochem. Zentralbl., 3, 568 (1905).
0.Schulz, "Untersuchungen uber die Wirking des Chinons und emiger Chinonderivate," Thesis,
Rostock, 1892.
S Cohn, "Ueber das Verhalten des Chinons im tierischen Organismus," Thesis, K6nigsberg,
1893.
F. Hoppe-Seyler, Z. Physiol. Chem., 7, 178 (1882).
A. Rymsza, "Em Beitrag zur Toxikologie der Pikrinsaure," Thesis, Dorpat, 1889.
J. P. Karplus, Z. Kim. Med., 22, 210(1893).
N. Sieber and A. Smirnow, Monatsh. Chem., 8, 88 (1887).
R. Cohn, Z Physiol. Chem., 17, 274 (1893).
R. Cohn, Ibid., 18, 133 (1894).
R. Kobert, Lehrbuch der Pharmakotherapie, Stuttgart, 1897, p.462.
0, Liebreich, Das Chloralhydrat, em neues Hypnoticum und Ana~stheticum, Berlin, 1869.
W. F. Loebisch, Die neueren Arzneimittel, 3rd ed., Vienna, 1888.
J. Personne, C. H. Acad. Sci. (Paris), 69, 979 (1869).
0. Hammarsten, Upsala Lakaretoren. Forehandl., 5, 424.
A. Rajewsky, Centralbl. Med. Wissensch., 8, 211 (1870).
E. Kulz, Sitzungsber. Ges. Befdrd. Naturwiss. (Marburg), 2, (1872).
M. Bouchut, C. R. Acad. Sci. (Paris), 69, 966 (1869).
A. Tomaszewicz, Pfluger's Arch. Physiol., 9, 35 (1874).
J. von Mering and F. Musculus, Ber. Dtsch. Chem. Ges., 8, 662 (1875).
C. P Falck, Dtsch. Z. Prakt. Med., 23, 64 (1877).
E. Kulz, Centralbl. Med. Wissensch., 19, 337 (1881).
0. Schmiedeberg and H. Meyer, Z. Physiol. Chem., 3, 422 (1879).
E. Kulz, Pfluger's Arch. Physiol., 28, 506(1882).
K. Garzarolli-Thurnlackh, Ann. Chem. Pharm., 210, 63 (1881).
J. von Mering, Ber. Dtsch. Chem. Ges., 15, 1019 (1882).
J. von Mering, Z. Physiol. Chem., 6, 480 (1882).
E. Kulz, Z. Biol., 20, 157 (1884).
50
[136]
[137]
[138]
[139]
[140]
[141]
[142]
[143]
[144]
CONTI AND BICKEL
0. L Erdmann, J. Prakt. Chem., ~33, 190 (1844); 37, 385 (1846).
W. Schmid, Ann. Chem. Pharm., 93, 83 (1855).
A. Baeyer, Ibid., 155, 257 (1870).
C A. Ewald, Centralbl. Med. Wissensch., Li, 819 (1873); Jahresber. Thierchem. , 3, 313 (1873).
J. von Mering, Centralbl. Med. Wissensch., 13, 945 (1875); Jahresber. Thierchem., 5, 61
(1875).
M. Jaffe, Z. Physiol. Chem., 2, 47(1878).
C. Wiedemann, Naunyn-Schmiedeberg's Arch. Exp. Pathol. Pharmakol., 6, 216 (1877).
M. Jaffe and R. Cohn, Ber. Dtsch. Chem. Ges., 20, 2311 (1887).
A Baeyer, Ibid., 10, 355 (1877).