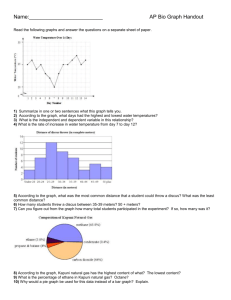
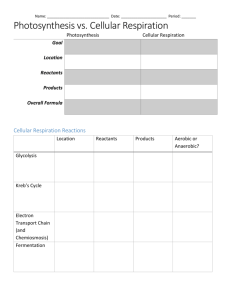
Chapter 21 Photosynthesis
advertisement

Chapter 21 Photosynthesis chloro1. Green 2. Chlorine (Cl) -plast living substance, organelle, cell Figure 21.1 Electron micrograph of a chloroplast. 21.1 Outline • What are the general properties of photosynthesis? • How is solar energy captured by chlorophyll? • What kinds of photosystems are used to capture light energy? • What is the molecular architecture of photosynthetic reaction centers? • What is the quantum yield of photosynthesis? • How does light drive the synthesis of ATP? • How is carbon dioxide used to make organic molecules? • How does photorespiration limit CO2 fixation? 21.2 The Sun - Ultimate Energy 1.5 x 1022 kJ of sunlight energy falls on the earth each day • 1% is absorbed by photosynthetic organisms and transformed into chemical energy • 6CO2 + 6H2O C6H12O6 + 6O2 • 1011 tons of CO2 are fixed globally per year • Formation of sugar from CO2 and water requires energy • Sunlight is the energy source! Figure 17.1 The flow of energy in the biosphere is coupled to the carbon and oxygen cycles. 21.3 21.1 What Are the General Properties of Photosynthesis? ['θailәˏkɔid] 葉綠層; 層狀體; 類囊膜 Grana 葉綠餅 基質 ['grænәm] [lә'melә] 薄層、片板 Figure 21.2 Schematic diagram of an idealized chloroplast. Chloroplasts possess DNA, RNA and ribosomes. 21.4 Photosynthesis Consists of Both Light Reactions and Dark Reactions thylakoid membranes reducing potential (NADPH) and ATP stroma The Calvin Cycle Hexose sugar Figure 21.4 The light-dependent and light-independent reactions of photosynthesis. 21.5 Water is the Ultimate e- Donor for Photosynthetic NADP+ Reduction The reaction sequence for photosynthesis in green plants: 2H2O + 2NADP+ + xADP + xPi → O2 + 2NADPH + 2H+ + xATP + xH2O A More Generalized Equation for Photosynthesis: CO2 + 2H2A → Hydrogen Hydrogen acceptor donor (CH2O) + 2A + Reduced Oxidized acceptor donor H2O In photosynthetic bacteria,H2A can be H2S or other oxidizable substrates, like isopropanol: Photosynthesis provides the oxygen on which we depend 21.6 21.2 How Is Solar Energy Captured by Chlorophyll? Figure 21.5 Structures (a) and absorption spectra (b) of chlorophyll a and b. The phytyl side chain of ring IV provides a hydrophobic tail to anchor the chlorophyll in membrane protein complexes. ['klorә,fiI] 葉綠素 Since the absorption spectra for a and b differ, plants that possess both can harvest a wider spectrum of incident energy. 21.7 Accessory Light-harvesting Pigments Increase the Possibility for Absorption of Light 藻藍膽素 [ˏfaikәu'saiәnɔbilin] Figure 21.6 Structures of representative accessory light-harvesting pigments in photosynthetic cells. (a) βCarotene, an accessory lightharvesting pigment in leaves. (b) Phycocyanobilin, a blue pigment found in cyanobacteria. 藍綠色的 21.8 The Light Energy Absorbed by Photosynthetic Pigments Has Several Possible Fates Figure 21.7 Possible fates of the quantum of light energy absorbed by photosynthetic pigments. Loss as heat Loss as light changes the reduction potential of the pigment 21.9 The Z scheme of photosynthesis Photosystem II splits water, producing O2, use quinones as terminal electron acceptors. Cyt b6f complex is a proton pump plastoquinone ['plæ stәukwinәun] 質體醌 Photosystem I provides reducing power, use photophosphorylation ferredoxins (Fd) as terminal electron acceptors. plastocyanin Fd: ferredoxins [ˏplæstә'saiәnin] 鐵氧化還原蛋白 質體藍素 Fp: flavoprotein, copper-containing ferredoxin-NADP+ reductase protein Figure 21.11 (b) The functional relationships among PSI, PSII, the cyt bf complex, and the ATP synthase in the thylakoid membrane. 21.10 The Z Scheme of Photosynthesis Ao: chlorophyll a A1: PSI quinone Figure 21.11 (a) The electron carriers are arranged according to their standard reduction potentials 21.11 Photosynthesis on Photosystems I (PSI) and PSII PSII splits water, producing O2, use quinones as terminal electron acceptors. 醌類 PSI provides reducing power, use ferredoxins as terminal electron acceptors. 鐵氧化還原蛋白 electrons flow from H2O to NADP+ Figure 21.10 Roles of the two photosystems, PSI and PSII. 21.12 Electrons Flow From Pheophytin in the Z Scheme Via Plastoquinones Figure 21.13 The structures of plastoquinone A and its reduced form, plastohydroquinone (or plastoquinol). Plastoquinone A has nine isoprene units and is the most abundant plastoquinone in plants and algae. Plastoquinone (PQ) exists as a pool within the chloroplast membrane. Because of its lipid nature, plastoquinone is mobile within the membrane and shuttles electrons from PSII to the cytochrome b6f complex. 21.13 Electrons from PSII Are Transferred to PSI via the Cytochrome b6f Complex • The cytochrome b6f complex is a large multimeric protein possessing 26 transmembrane α-helices • Cytochrome b6f complex is homologous to the cytochrome bc1 complex of mitochondria • The purpose of this complex is to mediate the transfer of electrons from PSII to PSI and to pump protons across the thylakoid membrane • Plastocyanin (PC) • a small copper-containing protein that carries electrons from cytochrome b6f to PSI • The copper in PC cycles between the reduced Cu+ and oxidized Cu2+ states in this transfer 21.14 21.4 What Is the Molecular Architecture of Photosynthetic Reaction Centers? 玫瑰紅色 Figure 21.14 The Rhodopseudomonas viridis reaction center (RC) is an integral membrane protein. (a) Diagram of RC showing light activation and path of e- transfer. Pheo: bacteriopheophytin QA: ubiquione A 21.15 Photosynthetic Electron Transfer by the R. viridis Center Leads to ATP Synthesis Figure 21.15 Photophosphorylation. Photoexcitation of R. viridis RC leads to reduction of a quinone, Q to form QH2. Oxidation of QH2 by the bc1 complex leads to H+ translocation for ATP synthesis. 21.16 How Does PSII Generate O2 From H2O? Figure 21.17 Structure of the PSII oxygen-evolving complex (OEC). Four Mn atoms (red, A-D) and a Ca atom (green) form the water-splitting metal cluster of the OEC. Bridging O atoms are purple. 21.17 The Architecture of PSI Resembles the R. viridis Center and also PSII Architecture Figure 21.18 The architecture of PSI. (a) Subunit organization. (b) A model for PSI. PsaA is orange; PgaB is magenta; PsaC is yellow; iron-sulfur clusters are red. 21.18 The Architecture of PSI Resembles the R. viridis Center and also PSII Architecture • PSI from S. elongatus is a cloverleaf-shaped trimer • Each monomer consists of 12 subunits and 127 cofactors • All of the prosthetic groups essential to function are located in just 3 of the subunits: PsaA, PsaB, PsaC • PsaA and PsaB form the reaction center heterodimer • PsaC interacts with the stromal face of the PsaA-PsaB dimer • The electron-transfer system of PSI consists of three pairs of chlorophyll molecules that mediate electron transfer to the quinone acceptor • The S. elongatus quinone is phylloquinone [ˏfilokwә`non] 葉綠醌, Vitamin K1, 凝血維生素一 21.19 How Do Green Plants Carry Out Photosynthesis? • Light-harvesting complex 1 (LHC1) is a supercomplex of 16 distinct protein subunits and 200 prosthetic groups • Four LHC1 subunits form an arc around one side of the PSI reaction center • A second LHC (LHC2) binds to another side • The many Chl molecules and other light-harvesting molecules of the supercomplex form an integrated network for highly efficient transfer of light energy into P700 Figure 21.19 View of the plant PSILHC1 supercomplex, from the stromal side of the membrane. Chlorophylls are green, lipids are red. 21.20 21.6 How Does Light Drive the Synthesis of ATP? • The transduction of the electrochemical gradient into the chemical energy of ATP is carried out by the chloroplast CF1CF0-ATP synthase • Protons pumped into the lumen of the thylakoids flow back out, driving the synthesis of ATP • CF1-CFo ATP synthase is similar to the mitochondrial ATP synthase 21.21 Cyclic Photophosphorylation Generates ATP but Not NADPH or O2 • Cyclic photophosphorylation depends only on PSI, not on PSII • It diverts the activated electron lost from PSI back through the PQ pool, the cytochrome b6f complex, and plastocyanin to rereduce P700+ Figure 21.21 The pathway of cyclic photophosphorylation by PSI. 21.22 Photosynthetic Electron Transport and ATP Synthesis Biology / Medicine Animations HD https://www.youtube.com/watch?v=LtecIPc30nM 21.23 Dark Reactions of Photosynthesis thylakoid membranes reducing potential (NADPH) and ATP stroma The Calvin Cycle Hexose sugar Figure 21.4 The light-dependent and light-independent reactions of photosynthesis. 21.24 21.7 How Is Carbon Dioxide Used to Make Organic Molecules? Fixation of CO2 is a unique ability of plants, algae, etc. • Melvin Calvin at Berkeley in 1945 showed that Chlorella could take up 14CO2 and produce 3phosphoglycerate • What was actually happening was that CO2 was combining with a 5-C sugar to form a 6-C intermediate • This breaks down to two 3-P-glycerates • Ribulose-1,5-bisphosphate is the CO2 acceptor in CO2 Fixation 21.25 Ribulose-1,5-Bisphosphate is the CO2 Acceptor in CO2 Fixation RuBP carboxylase 二磷酸核酮糖羧化酶 (Rubisco) Ribulose-1,5Bisphosphate (RuBP) Figure 21.23 The ribulose bisphosphate carboxylase reaction. Mg2+ at the active site aids in stabilizing the 2,3-enediol transition state (I) for CO2 addition and in facilitating the carbon-carbon bond cleavage that leads to product formation. 21.26 Ribulose-1,5-Bisphosphate is the CO2 Acceptor in CO2 Fixation 3-P-glycerates Figure 21.23 The ribulose bisphosphate carboxylase reaction. Mg2+ at the active site aids in stabilizing the 2,3-enediol transition state (I) for CO2 addition and in facilitating the carbon-carbon bond cleavage that leads to product formation. 21.27 The Calvin-Benson Cycle (The Calvin Cycle ) • The set of reactions that transform 3-P-glycerate into hexose sugar • The principal CO2 fixation pathway in nature • A large part of this pathway is really a disguised gluconeogenesis pathway! • With some pentose phosphate pathway reactions thrown in 21.28 CO2 Fixation into Carbohydrate Proceeds Via the Calvin-Benson Cycle Net: 6 CO2 + 18 ATP + 12 NADPH + 12 H+ + 12 H2O --> glucose + 18 ADP + 18 Pi + 12 NADP+ 21.29 CO2 Fixation into Carbohydrate Proceeds Via the Calvin-Benson Cycle (Rubisco) 21.30 CO2 Fixation into Carbohydrate Proceeds Via the Calvin-Benson Cycle 21.31 The Carbon Dioxide Fixation Pathway Is Indirectly Activated by Light • The activities of key Calvin cycle enzymes are coordinated with the output of photosynthesis • In effect, these enzymes respond indirectly to light activation • Light induces pH changes in chloroplast compartments • Rubisco, rubisco activase, and several Calvin cycle enzymes are more active at alkaline pH • Light energy generates reducing power • Reduced ferredoxin and NADPH • Light induces movement of Mg2+ ions from the thylakoid vesicles into the stroma 21.32 The Carbon Dioxide Fixation Pathway Is Indirectly Activated by Light Figure 21.25 Light regulation of CO2 fixation prevents a substrate cycle between cellular respiration and hexose synthesis by CO2 fixation. 21.33 The Carbon Dioxide Fixation Pathway Is Indirectly Activated by Light Figure 21.26 Light-induced pH changes in chloroplast compartments. These pH changes modulate the activity of key Calvin cycle enzymes. 21.34 21.9 How Does Photorespiration Limit CO2 Fixation? • Ribulose bisphosphate carboxylase/oxygenase catalyzes an alternative reaction in which O2 replaces CO2 as the substrate added to RuBP • This oxygenase reaction diminishes plant productivity because it leads to loss of RuBP, the CO2 acceptor • The products of the oxygenase reaction are 3phosphoglycolate and phosphoglycolate • Dephosphorylation and oxidation convert phosphoglycolate to glyoxylate • Transamination yields glycine 21.35 Photosynthesis (Dark Reactions, Calvin Cycles) Biology / Medicine Animations HD https://www.youtube.com/watch?v=joZ1EsA5_NY 21.36 The Hatch-Slack Pathway: The C-4 Pathway for CO2 Fixation 葉肉細胞 束鞘細胞 Figure 21.29 Essential features of the compartmentation and biochemistry of the Hatch-Slack pathway of carbon dioxide uptake in C4 plants. 21.37