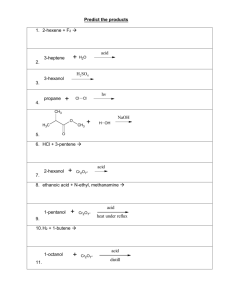
Coenzymes, vitamins, trace elements
advertisement

Coenzymes, vitamins, trace elements Josef Fontana EC - 41 Overview of the lecture • Introduction to the topic: cofactors – Revision of the enzyme structure – The interrelationship cofactor - vitamin - trace element • Simple enzymes (proteases) • Examples of complex enzymes and function of cofactors in their molecules – Vitamins as cofactors (decarboxylases, dehydrogenases) – Trace elements as cofactors (carbonic anhydrase) – Cofactors of enzyme groups • The importance for doctors - deficiency as a disease / deficiency as a treatment of diseases Introduction to the topic: cofactors Revision of the enzyme structure Structure and properties • most of enzymes are proteins The figure is found at: http://fig.cox.miami.edu/~cmallery/255/255enz/enzymology.htm (December 2006) Enzyme structure • Enzymes are mostly proteins (exception: ribozyme catalytic active RNA). • Some enzymes in addition to protein component contain also non-protein component. According to this we can divide enzymes into: – Simple enzymes contain only protein (pepsin, trypsin, ribonuclease). – Complex enzymes contain protein and non-protein component = cofactor. • Cofactor is the non-protein part of the enzyme molecule. It is necessary for its catalytic function. Our body can not synthesize them often - therefore, we eat their precursors – e.g. vitamins. Introduction to the topic: cofactors The interrelationship cofactor - vitamin - trace element Cofactor can be • 1) metal ion: Zn2+, Mn2+, Mg2+, Fe2+, Cu2+ (trace elements) • 2) organic molecule: – coenzymes are slightly bound to the enzyme, undergo a chemical change and are released from the enzyme molecule, they are derivates of vitamins very often: NAD(P)+, FAD, coenzyme Q, ... – prosthetic groups are tightly bound to the enzyme and remain associated with enzyme during the whole reaction: heme, FAD,… • Coenzyme + apoenzyme (inactive protein) → holoenzyme (active enzyme) The figure is found at: http://stallion.abac.peachnet.edu/sm/kmccrae/BIOL2050/Ch1-13/JpegArt1-13/05jpeg/05_jpeg_HTML/index.htm (December 2006) Simple enzymes (proteases) Proteases - enzymes that cleave the peptide bond H O R' H N N N R H O H H2O protease R' O N OH R + H N H2N O Bond cleavage requires the presence of water (hydrolysis). Side chains of amino acids play a key role in the reaction Simple enzymes • How looks like the catalytic mechanism of simple enzymes ? I show you two examples: • 1) pepsin, which belongs to the aspartate proteases and • 2) chymotrypsin, which belongs to the serine proteases • Neither of them needs to have cofactor for the hydrolysis. Both need properly arranged side chains of amino acids in their active site. How works pepsin • For the pepsin catalytic activity are essential two aspartates (their side chains) in its active site and water molecule. • One aspartate attracts proton from water molecule. Water then attacks the carbonyl group (that forms a peptide bond). How works pepsin • Catalysis involves two conserved aspartyl residues. • In the first stage of the reaction, an aspartate functioning as a general base (Asp X) extracts a proton from a water molecule, making it more nucleophilic. • This resulting nucleophile then attacks the electrophilic carbonyl carbon of the peptide bond targeted for hydrolysis, forming a tetrahedral transition state intermediate. How works pepsin • A second aspartate (Asp Y) then facilitates the decomposition of this tetrahedral intermediate by donating a proton to the amino group produced by rupture of the peptide bond. • The two different active site aspartates can act simultaneously as a general base or as a general acid because their immediate environment favors ionization of one, but not the other. How works chymotrypsin • While catalysis by aspartic proteases involves the direct hydrolytic attack of water on a peptide bond, catalysis by the serine protease chymotrypsin involves prior formation of a covalent acyl enzyme intermediate. • A highly reactive seryl residue, serine 195, participates in a charge-relay network with histidine 57 and aspartate 102. • Far apart in primary structure, in the active site these residues are within bond-forming distance of one another. Aligned in the order Asp 102-His 57-Ser 195, they constitute a "charge-relay network" that functions as a "proton shuttle." How works chymotrypsin CH2OH H N CH2 N CH2-COOH all these tools come from amino acids in the protein active site How works chymotrypsin • Binding of substrate initiates proton shifts that in effect transfer the hydroxyl proton of Ser 195 to Asp 102. • The enhanced nucleophilicity of the seryl oxygen facilitates its attack on the carbonyl carbon of the peptide bond of the substrate, forming transient tetrahedral intermediate. How works chymotrypsin • The proton on Asp 102 then shuttles through His 57 to the amino group liberated when the peptide bond is cleaved. • The portion of the original peptide with a free amino group then leaves the active site and is replaced by a water molecule, yielding an acyl-Ser 195 intermediate (covalent bond acyl-enzyme). How works chymotrypsin • The charge-relay network now activates the water molecule by withdrawing a proton through His 57 to Asp 102. • The resulting hydroxide ion attacks the acyl-enzyme intermediate, forming a second tetrahedral intermediate. • The charge-relay system donates a proton to Ser 195 (restoring its original state), facilitating breakdown of tetrahedral intermediate to release the carboxyl terminal peptide. Examples of complex enzymes and function of cofactors in their molecules Vitamins as cofactors (decarboxylases, dehydrogenases) Vitamins • Name derives from the Latin word Vita = Life • Vitamins soluble in water: • Vitamin C • B complex: B1, B2, B3, B6, folic acid (B9), B12, niacin (PP), biotin, pantothenoic acid • Fat-soluble vitamins: Vitamin A, D, E, K B1 Thiamine • Active cofactor is thiamine pyrophosphate - TPP (produced by TPPsynthetase) • TPP is a cofactor in oxidative decarboxylation of pyruvate and alphaketoglutarate. • In addition, TPP is used in transketolase reactions (pentose phosphate pathway). Thiamine → TPP Thiamine pyrophosphate • The key part of the cofactor molecule is thiazole ring with its acidic hydrogen. • Hydrogen is removed by the enzyme, forming carbanion. Molecule is called ylide (contains anion next to the cation). • Anion can then react with the carbonyl group in different molecules (e.g. pyruvate). • Pyrophosphate works as a handle, which holds the cofactor in proper place within the enzyme molecule. Thiamine pyrophosphate • The key part of the cofactor molecule is thiazole ring with its acidic hydrogen. • Hydrogen is removed by the enzyme, forming carbanion. Molecule is called ylide (contains anion next to the cation). Thiamine pyrophosphate • TPP catalyses the cleavage of a substrate compound at a carbon-carbon bond connecting a carbonyl group to an adjacent reactive group (usually a carboxylic acid or an alcohol). • It achieves this in four basic steps. O Cl Cl CH3 Thiamine pyrophosphate N O- N O H • 1) The carbanion of the TPP ylid nucleophilically attacks the carbonyl group on the substrate. • This forms a single bond between the TPP and the substrate. CH3 H pyruvate S S ylid acidic hydrogen Cl CH3 Cl - CO2 CH3 N N S HO S O OH O- resonance Cl CH3 CH3 N N H + S S OH O H H O + H3C H acetaldehyde ylid O Cl Cl CH3 Thiamine pyrophosphate N O- N O H • 2) The target bond on the substrate is broken, and its electrons are pushed towards the TPP. • This creates a double bond between the substrate carbon and the TPP carbon and pushes the electrons in the N-C double bond in TPP entirely onto the nitrogen atom, reducing it from a positive to neutral form. CH3 H pyruvate S S ylid acidic hydrogen Cl CH3 Cl - CO2 CH3 N N S HO S O OH O- resonance Cl CH3 CH3 N N H + S S OH O H H O + H3C H acetaldehyde ylid O Cl Cl CH3 Thiamine pyrophosphate N • 4) The TPP-substrate bond is broken, reforming the TPP ylid and the substrate carbonyl (decarboxylated substrate) is transferred on the target molecule (e.g. lipoamide). O- N O H • 3) The electrons push back in the opposite direction forming a new bond between the substrate carbon and hydrogen proton. CH3 H pyruvate S S ylid acidic hydrogen Cl CH3 Cl - CO2 CH3 N N S HO S O OH O- resonance Cl CH3 CH3 N N H + S S OH O H H O + H3C H acetaldehyde ylid thiazolium ring NH2 H C N N S O H3C N H3C H2 C H2C O O P O P O- O- O- thiamine pyrophosphate H NH2 H3C C OH C N N S O H3C N H3C H2C H2 C O P O- Hydroxyethyl thiamine pyrophosphate O O P O- O- Lipoic acid • Lipoic acid is a co-factor found in pyruvate dehydrogenase and α-ketoglutarate dehydrogenase and two multienzymes involved in α-keto acid oxidation. • Lipoic acid couples acyl group transfer and electron transfer during oxidation and decarboxylation of α-ketoacids. • No evidence exists of a dietary lipoic acid requirement in humans. Therefore it is not considered to be a vitamin. S SH HS S CH H2C COOH C H2 lipoic acid, reduced form H S CH H2C C H2 COOH C H2 lipoic acid, oxidized form S CH H2C NH N C O CH C O lipoamide complex (lipoyl-lysine conjugate) Lipoic acid exists in 2 forms: a closed-ring disulfide form and an open-chain reduced form; oxidation-reduction cycles interconvert these 2 species; lipoic acid exists covalently attached in an amide linkage with lysine residues on enzymes B2 Riboflavin • It forms 2 cofactors : • flavin mononucleotide, FMN • flavin adenine dinucleotide, FAD • Involved in the metabolism of carbohydrates, fats and proteins (flavin dehydrogenases / flavoproteins). • Hydrogen carriers in the respiratory chain H3C H2C OH H C OH H C OH H C OH H C H N N O N H3C N H O RIBOFLAVIN Riboflavin H H3 C N N O N H O FAD (oxidized form) N N O N N H3 C H3C H3C N H hydrogen addition occurs in 2 steps H O FADH2 (reduced form) Succinate dehydrogenase CO2CH2 succinate C H CH2 CO2- CO2- succinate dehydrogenase FAD FADH2 H C CO2fumarate Xanthine oxidase OH OH N N OH N N N N OH N N HO N N H hypoxanthine HO N N H xanthine H uric acid xanthine oxidase Xanthine oxidase is a flavoprotein which also contains Fe and Mo – trace elements Examples of complex enzymes and function of cofactors in their molecules Trace elements as cofactors (carbonic anhydrase) Trace elements • Present in human body in amount less than 1mg/kg. • I, Zn, Cu, Se, Cr, F, Mn, Co, Mo • Have known biological function (prosthetic groups). • Deficiencies are huge problem in developing countries. • But also specific risk groups: the homeless, alcoholics and patients on total parenteral nutrition (TPN) Carbonic anhydrase • Catalyzes the reaction: • The active site contains zinc. Carbonic anhydrase • A zinc prosthetic group in the enzyme is held by 3 histidine side-chains. The fourth coordination position is occupied by water. • This causes polarisation of the hydrogen-oxygen bond in water, making the oxygen slightly more negative, thereby weakening the bond. Carbonic anhydrase • A fourth histidine is placed close to the substrate of water and accepts a proton. This leaves a hydroxide attached to the zinc. • The active site also contains specificity pocket for carbon dioxide, bringing it close to the hydroxide group. This allows the electron rich hydroxide to attack the carbon dioxide, forming bicarbonate. • Bicarbonate is replaced with a new water molecule. Examples of complex enzymes and function of cofactors in their molecules Cofactors of enzyme groups Cofactors help to catalyze many reactions • Cofactors of oxidoreductases: NAD(P)+, FAD, cytochromes (contain heme), Fe-S complexes • Coenzymes carrying C1 radicals: tetrahydrofolate, vitamin B12, Sadenosylmethionine, biotin (cofactor of carboxylases) • Cofactors carrying acyl: lipoic acid (PDH prosthetic group, α-KGDH) HSCoA, pyridoxal phosphate (transaminases) Cofactors of oxidoreductases NAD+ NADP+ nicotinamide adenine dinucleotide nicotinamide aden. dinucl. phosphate (precursor: niacin = nicotinic acid) H+ FAD FMN flavin adenine dinucleotide flavin mononucleotide 2 H+ (precurzor: riboflavin = vitamin B2) heme Fe3+ + e- ↔ Fe2+ ⇒ e- Cofactors of transferases ATP GTP TDP adenosine triphosphate guanosine triphosphate thiamine diphosphate / phosphate / phosphate / C-fragment PALP pyridoxal phosphate / -NH2 (prekurzor: thiamine = vitamin B1) (prekurzor: pyridoxine = vitamin B6) THF tetrahydrofolate (prekurzor: folic acid) / C1-fragment CoA coenzyme A (HS-Co-A) / acyl PAPS phosphoadenosine phosphosulfate / sulfate Cofactors of Lyases: PALP pyridoxal phosphate (decarboxylases) Ligases: ATP adenosine triphosphate → acyl-CoA-synthetases → aminoacyl-tRNA-synthetases biotin = vitamin H (carboxylases) Coenzymes and prosthetic group NAD+ ↔ NADH + H+ nicotinamide adenine dinucleotide coenzyme FAD ↔ FADH2 flavin adenine dinucleotide (vit. B2 = riboflavin) prosthetic gr. Other examples: coenzyme A, coenzyme Q, tetrahydrofolate, thiamine diphosphate (vit. B1 = thiamine) http://web.indstate.edu/thcme/mwking/vitamins.html Prosthetic groups Biotin (vit. H) Heme • Another example: pyridoxal phosphate (derivate of vitamin B6) http://web.indstate.edu/thcme/mwking/vitamins.html The importance for doctors deficiency as a disease / deficiency as a treatment of diseases B9 Folic Acid • Folic acid is composed of pteridine, P-amino benzoic acid, and glutamic acid. • Its derivate is tetrahydrofolate. It can carry one-carbon fragments (involved in the methylation reactions). Folate deficiency causes megaloblastic anemia • N5, N10-methylene-tetrahydrofolate carries methyl group into the thymine synthesis (necessary for DNA synthesis and thus also for the formation of erythrocytes). • Deficiency of folic acid affects cells that are dividing rapidly because they have a large requirement for thymidine for DNA synthesis. • Clinically, this affects the bone marrow, leading to megaloblastic anemia. Lack of folate can treat tumors • During the thimine synthesis there are several changes in the N5, N10-methylene-THF molecule. • The methylene group is reduced to the methyl group that is transferred, and tetrahydrofolate is oxidized to dihydrofolate. • For further pyrimidine synthesis to occur, dihydrofolate must be reduced back to tetrahydrofolate, a reaction catalyzed by dihydrofolate reductase. • Dividing cells, which must generate thymidine and dihydrofolate, thus are especially sensitive to inhibitors of dihydrofolate reductase such as the anticancer drug methotrexate. Lack of folate can treat infections • Human body can not produce its own THF, while pathogenic bacteria have this ability. • This is used in the treatment of bacterial diseases with sulphonamides. • They are in fact for bacteria indistinguishable similar to paminobenzoic acid. • Bacteria try to incorporate sulphonamides into THF molecules, but the result is a dysfunctional cofactor. • Sulfonamides are the competitive inhibitors of enzymes involved in synthesis of THF.