brief description
advertisement

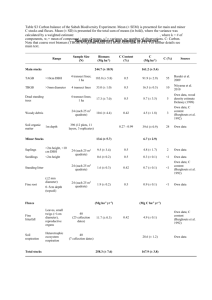
Gentry 0.1 ha transect methodology, as modified by Brad Boyle (excerpt from Boyle 1996, Chap. 1). Modified 0.1 ha sampling methodology. The basic sampling unit was a census of woody plants 2.5 cm dbh (diameter at breast height) within a 0.1 ha linear transect. Each had the same discontinuous form of five parallel 2 m x 100 m subunits, laid out with 10 m separation between the midline of each subunit. In practice, a single 0.1 ha census was done 50 m at time, with each 50 m segment being referred to as a "line" (Fig. 1). Although the direction for the initial line was chosen to run roughly perpendicular to the slope, subsequent lines followed the initial compass direction, without attempting to conform to elevational contours. A 50 m tape marked the middle of the long axis of each line; individual plants were censused if they lay within 1 meter of either side of the tape (subject to the rules discussed below). Slope was recorded every 10 meters along each line with a hand-held clinometer, and the average of these measurement was used as the single slope value for each 0.1 ha sample. All woody growth forms—trees, shrubs, lianas and hemiepiphytes—were included in the census. Inclusion criteria differed according to growth habit. Trees and shrubs were counted if the center of the base of the trunk lay within one meter of either side of the tape and the dbh (diameter at breast height) was >= 2.5 cm. Dbh was defined as 1.3 m from the base of the trunk—not necessarily above the ground; thus, live, rooted trees pressed flat to the bround (e.g., by branch falls) were also included if their trunks were of sufficient thickness at 1.3 m from the base. For hemiepiphytes, a plant was included if any part of the aerial root (in the case of primary hemiepiphytes, such as Clusia or Ficus) or rhizome (for secondary hemiepiphytes, such as most Araceae) had a diameter of >= 2.5 cm at or below 1.3 m above ground level, within the horizontal boundaries of the line. Lianas were counted if one or more of their roots entered the soil anywhere within the horizontal limits of the line (i.e., within 1 m either side of the midline) and if they had a maximum diameter of 2.5 cm at any observable point along the stem, inside or outside of the 2 m boundary. Stems were tallied as separate individuals if they were not obviously connected above the ground. This undoubtedly overestimates the number of individuals within a transect, but avoids counting as "individuals" the frequent instances of multiple above-ground divisions of a single trunk. Basal area can still be calculated for a given individual by summing the basal areas of its constituent stems. Differences from the Gentry methodology. The method just described is nearly identical to the 0.1 ha transect method of Gentry (Gentry 1982, 1988). The main difference is in the regular layout of the 2 x 50 m lines. In the Gentry method, the lines are laid out one after the other without attempting to maintain a set compass direction. I adopted the more regular layout for three reasons. One was to reduce the observer bias inherent in the choice of location for each line. With the method used in this study, the only element of subjectivity enters into the inital selection of a starting point and compass direction for the first line; subsequent lines must conform to the predetermined layout, regardless of terrain or forest composition. The second reason was to reduce the confounding of local diversity with beta diversity caused by traversing different microhabitats, an inherent problem of long, narrow samples. The third reason was to ensure repeatability and comparability among samples, an obvious pre-requisite for statistical comparisons. Although the precise dimensions of the transect -- for example, the distance between the parallel 100 m subunits -- was somewhat arbitrary, the motivation behind the use of discrete segments was to subsample a larger, uniformly-shaped area of forest (in this case, a 40 m x 100 m rectangle, or roughly 0.5 ha), thereby reducing the between-sample variance due to clumping and/or hyper-dispersion among individuals. 1 Literature cited Boyle, B. L. 1996. Changes on altitudinal and latitudinal gradients in neotropical montane forests. Washington University, St. Louis. 275 pp. Gentry, A. H. 1982. Patterns of neotropical plant species diversity. Evolutionary Biology. Hecht, Wallace and Prance, Plenum Publishing Corporation. 15: 1-84. Gentry, A. H. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75: 1-34. Fig. 1. Layout of a single 0.1 ha transect, as modified by Boyle (1996). Each line is censused by laying out a 50 m tape and counting all plants within 1 m of either side, according to the inclusion criteria discussed in text. Length of each 50 m line is arbitrary; sample can also be done as five parallel lines if a 100 m tape is available. 2