Exercise Metabolism:
advertisement

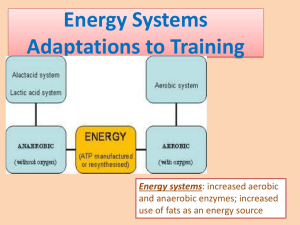
Lab IV Part 1: Exercise Metabolism: Fuel substrate selection Part 2: Thermoregulation & exercise Introduction Exercise is one of the greatest energy stresses that an organism is likely to encounter. Like other stresses, severe exercise has the potential to disrupt homeostasis. Every cell requires a certain amount of energy for life-sustaining processes. Thus, any disruption of cellular energy homeostasis is potentially fatal for the cell. Fortunately, we have evolved several potent mechanisms to maintain cellular energy homeostasis. Nearly every change in our body's physiology during exercise is centered on maintaining cellular energy homeostasis. For example, the increases in ventilation and cardiac output are meant to increase oxygen delivery to the mitochondria in active tissues, where oxygen serves as the final electron acceptor in the electron transport chain. The importance of energy metabolism during exercise is also illustrated by the fact that many of the possible causes of fatigue are related to cellular energy metabolism (lab 4). The energy that allows us to perform life-sustaining processes, accomplish work, grow, and maintain body temperature ultimately comes from our food stuffs. However, the energy contained in our food stuffs is not readily accessible as a source of energy for the energy requiring processes of the cell. The energy from our food must first be converted to a common energy currency; ATP (Adenosine triphosphate). It is ATP that serves as the immediate energy source for muscular work. Because muscle ATP concentrations are very low, the production of ATP, sometimes at very high rates, is of the highest priority during exercise. If exercise physiology were a story, the plot would center on energy and work. It is therefore extremely important for exercise physiology students to understand the basic principals and concepts related to exercise metabolism. By this time students have already learned in their introductory biology, human physiology, and exercise physiology courses about the following basic cellular metabolic processes: the ATP-PC system, glycolysis, glycogenolysis, gluconeogenesis, lipolysis, beta oxidation, the tricarboxylic acid cycle, and the electron transport chain. It is recommended that students review these processes before coming to lab. These processes will only be discussed briefly in order to help students understand the “big picture” of exercise metabolism; a complete review of these processes is beyond the scope of this lab. Today's laboratory experiments are meant to help students better understand those factors (e.g. exercise intensity, duration, etc) that determine what fuel sources and metabolic pathways are contributing to an exercise bout. Exercise as a metabolic stress At rest, almost all of our energy is coming from aerobic metabolism, and results in an oxygen consumption of around 3.5 ml/kg/min, or about 0.25 L/min in an average human. Assuming a caloric equivalent of 5 kcal/L, this means that, at rest most of us expend about 1 to 1.5 kcal/minute. An average college age male can reach energy expenditures of 15-20 kcal/minute at maximal exercise. Olympic rowers average 36 kcal/min during a simulated 2000m race lasting 57 minutes, and can attain energy expenditures up to 40 kcal/min during training and competition – a 40-fold increase in energy expenditure above rest! Of course these high rates of energy expenditure can not be sustained for long periods of time. Among the highest measured daily energy budgets to be maintained for more than a few days were obtained from cyclists participating in the Tour de France. In these cyclists daily energy expenditure was around 7,000 kcal/day for 22 days. This is almost 4.5 times the typical basal metabolic rate (~1,600 kcal/day) for these athletes. Exercise clearly places extreme demands on the body’s energy systems. For example, it has been demonstrated that flux through the tricarboxylic acid cycle (also called the citric acid cycle or Krebs cycle) can increase over 100-fold during moderate to high intensity exercise. Lab IV - 1 Fuel substrates & stores in the human body Overall, the body has about 180,000 kcal of stored energy of which 70-80% (~140,000 kcal) are fats, ~20-30% are proteins (about 40,000 kcal), and less than 1% carbohydrates (usually < 1,500 kcal). The contribution of these various fuel sources to exercise energy expenditure is highly dependent upon diet, training, exercise intensity, and exercise duration. Most of body's energy stores are in the form of fats. Triglycerides (TG) are the primary storage form of fats in the body. Almost all of the body's TG stores are found in adipose tissue, but a small amount (2-3%) of the fat stores are found within skeletal muscle (intramuscular TG). Proteins are the next largest reserve of calories in the body. However, protein catabolism generally contributes very little to exercise energy expenditure (typically less than 2% of all energy expended, although it can reach 5-15% of energy expended during exercise lasting over 34 hours). Proteins can, on the other hand, play an important role as a "reserve" of energy stores during prolonged fasting. The body also stores carbohydrates in the form of glycogen. Most of the body's glycogen stores are in the muscles, with a smaller amount of glycogen stored in the liver. While small, the liver glycogen stores are very important for maintaining blood glucose concentrations between meals; glucose liberated from glycogen breakdown in the liver can enter the circulation. The breakdown of glycogen is called glycogenolysis, and is catalyzed by the enzyme phosphorylase. The making of glycogen from glucose and is called glycogenesis, and the final step of this process is catalyzed by the enzyme glycogen synthase. Certain tissues, for example nervous tissue, are very dependent upon glucose from the circulation. The maintenance of blood glucose is therefore extremely important at rest and during exercise. How can we maintain blood glucose with such limited supplies of carbohydrates? First, we consume carbohydrates on a regular basis, and second, certain tissues in the body (liver, kidney) are able to make glucose out of non-carbohydrate precursors (the making of glucose from amino acids, glycerol, and lactate is called gluconeogenesis). Liver glycogen stores are used primarily to maintain blood glucose concentrations between meals or when they are absolutely necessary (e.g. exercise). The regulation of blood glucose and glycogen breakdown is coordinated by the endocrine system. Several hormones can stimulate glycogen breakdown and increase blood glucose while insulin, on the other hand, can decrease blood glucose and stimulate storage of glycogen. In summary, the liver plays a central role in regulating blood glucose – it stores a small amount of glycogen that can be broken down to increase blood glucose when necessary, and has the ability to make glucose by gluconeogenesis. This regulation is mediated by several hormones. The skeletal muscle glycogen stores are larger than liver glycogen stores, so why doesn't skeletal muscle glycogen play a central role in regulation of blood glucose? In order to answer this question, students must first understand a few things about glucose metabolism in the cell and glucose transport into and out of the cell. Glucose is transported across the cell membrane by glucose transporters, called GLUTs. In skeletal muscle, the GLUT that is involved in both exercise-stimulated and insulin-stimulated glucose uptake is GLUT4. Once glucose enters a cell, it is rapidly phosphorylated (a phosphate group is attached to it), making glucose-6-phosphate. GLUT molecules are unable to transport glucose-6-phosphate. Liver cells have an enzyme (glucose-6-phosphatase) that can remove this phosphate group, but skeletal muscle cells do not. Thus, once glucose enters a skeletal muscle cell it can not leave. In summary, when glycogen is broken down in the liver, glucose can re-enter the circulation, but when glycogen is broken down in muscle, the resulting glucose can not re-enter the circulation. Whereas the primary role of liver glycogen is the maintenance of blood glucose, the primary role of skeletal muscle glycogen is to serve as an immediate source of energy during muscle activity. In fact, carbohydrate metabolism during moderate or high intensity exercise relies more on muscle glycogen as a substrate than glucose (from the circulation). Muscle glycogen depletion is a possible cause of fatigue during exercise lasting over 90 minutes. Thus, sparing muscle glycogen would benefit individuals during endurance exercise. Lab IV - 2 Exercise training tends to increase the use of fatty acids as a fuel source, thus at any given intensity a trained individual has to rely less on glycogen; sparing their glycogen stores. There is some evidence that caffeine may spare glycogen stores by increasing reliance on fat metabolism. However, there is also evidence that caffeine has no effect on sparing muscle glycogen. Carbohydrate ingestion during exercise is another strategy that has been used to help spare glycogen (by providing an alternative source of glucose for glycolysis). Blood glucose concentrations are usually well maintained during exercise, but during prolonged exercise lasting hours, blood glucose may eventually drop. Carbohydrate ingestion would also help avoid an undesired drop in blood glucose. The fatty acids used to produce energy by skeletal muscle are derived primarily from adipose tissue triglycerides, and to a lesser extent from intramuscular triglycerides. It should be noted that triglycerides (TG) must be broken down before they can be used as a fuel substrate. The degradation of TG into fatty acids and glycerol is called lipolysis. Inside the cells, lipolysis is catalyzed by the enzyme hormone sensitive lipase (HSL). A different lipase, lipoprotein lipase (LPL), catalyzes the breakdown of TG in the circulation and allows adipose cells to take up fatty acids from these circulating TG. Once fatty acids have been taken up by adipose cells they can be stored as TG, this is called lipogenesis. While both LPL and HSL are involved in triglyceride breakdown, they play very different roles in fat metabolism. LPL tends to increase storage of fatty acids in adipose tissue, whereas HSL mobilizes fatty acids so they can be used as an energy source by other tissues. Summary of the storage and breakdown of carbohydrate and lipid fuel stores: Glycogenolysis, catalyzed by phosphorylase Glycogen Glucose Glycogenesis, catalyzed by glycogen synthase Lipolysis, catalyzed by hormone sensitive lipase & lipoprotein lipase Free fatty acids & glycerol Triglycerides Lipogenesis, catalyzed by several enzymes Energy Systems and Metabolic Pathways As previously stated, ATP is the immediate source of energy for muscular activity, but is stored in only low concentrations in the cell. Thus, ATP must be rapidly resynthesized during exercise. There are multiple pathways involved in the production of ATP. The simplest and fastest mechanism of ATP production is the ATP-PC system (also called the phosphagen or phosphocreatine system). Phosphocreatine (usually abbreviated PC or PCr) is a high energy compound that can readily "donate" its phosphate group to ADP in order to rapidly produce ATP. This reaction, which is catalyzed by the enzyme creatine kinase, is summarized below. ADP & PCr Creatine Kinase ATP + Cr This reaction is reversible and does not require oxygen. During exercise, when ATP is being used rapidly and ADP concentrations increase, this reaction favors production of ATP at the expense of PCr. During recovery, the PCr stores must be replenished (which, of course requires ATP). The ATP-PC system is used at the beginning of any exercise bout, and because it can produce ATP so quickly it is especially important for high intensity exercise lasting less than 10 seconds in duration (see figure 1). Conversely, during long duration exercise, the contribution of the ATP-PC system to overall energy production is relatively small. Glycolysis is the breakdown of six-carbon glucose molecules into two three-carbon pyruvate molecules in the cytoplasm of the cell. Pyruvate produced during glycolysis has two Lab IV - 3 possible fates. One fate of pyruvate, which requires oxygen results in the formation of acetyl CoA, which can then enter the citric acid cycle. The other major fate of pyruvate in exercising skeletal muscle, which does not require oxygen, is the formation of lactate. Glycogen Glucose Aerobic Glycolysis Pyruvate Anaerobic Mitochondria Citric Acid Acetyl CoA Cycle & Electron Transport Chain Lactate Figure 2. Highly simplified summary of the aerobic and anaerobic metabolism of carbohydrates Thus, glycolysis concludes with either 1) the formation of lactate, which does not require oxygen, or 2) the formation of acetyl CoA under aerobic conditions. Therefore, glycolysis is sometimes referred to as aerobic or anaerobic glycolysis depending on which product is formed; acetyl CoA or lactate. Many students tend to think of lactate as just a by-product of anaerobic metabolism, but it is not just a by-product. Lactate formed in skeletal muscle can enter the circulation, be taken up by the liver, and converted back to glucose via gluconeogenesis. This glucose can then re-enter the circulation and be returned to the muscle, which can use it as a fuel substrate. This cycle (muscle lactate liver glucose muscle) is called the Cori cycle. The anaerobic energy systems include the ATP-PC system and anaerobic glycolysis. The term anaerobic means that these systems do not require oxygen. It is a common student misconception that these systems are only used when the cells are lacking oxygen. This is false. It is true that if a cell lacks oxygen it will have to rely on anaerobic energy systems to produce ATP. However, most of the cells in our body typically are able to maintain oxygen concentrations high enough for normal mitochondrial function; even during high intensity exercise. If the cells have sufficient oxygen for aerobic metabolism, why are anaerobic energy systems called upon to create ATP during high intensity exercise? The answer is that during high intensity exercise aerobic energy systems can not produce ATP fast enough to meet the muscle cell’s very high ATP demand. The anaerobic energy systems, which can produce ATP at high rates, are therefore necessary to meet the cell's ATP demands. A well known exercise physiologist, George Brooks, suggests that aerobic glycolysis be referred to as slow glycolysis and anaerobic glycolysis be referred to as fast glycolysis, in order to avoid this confusion. ATP-PC Rate of ATP resynthesis during maximal exercise Anaerobic Glycolysis Oxidative Metabolism 0 10sec 30 sec 60sec 90 sec 180 sec Duration Figure 1. Relative role of the different energy systems in making ATP during maximal exercise of different durations. Lab IV - 4 Hours Anaerobic (fast) glycolysis is much faster than aerobic glycolysis partly owing to the fact that it is a simpler process (it requires fewer steps to produce ATP) and aerobic glycolysis is slower partly because the production of ATP from acetyl CoA requires the citric acid cycle and the electron transport chain. It should, however, be noted that while anaerobic production of ATP during glycolysis is much faster, the total amount of ATP produced from each molecule of glucose is far greater during aerobic than during anaerobic glycolysis. It should also be noted that while the anaerobic energy systems can produce ATP fairly quickly, they can not maintain these high rates of ATP production for very long; sustained ATP production by the muscle cells requires aerobic ATP production. Before we discuss aerobic metabolism in greater detail we should recap anaerobic energy systems. The two anaerobic energy systems are the ATP-PC system and anaerobic glycolysis. The ATP-PC system is the fastest mechanism our cells have for producing ATP and is especially important for very high intensity, short duration exercise. Anaerobic glycolysis does not produce ATP as fast as the ATP-PC system, but is substantially faster than aerobic energy systems. As you can see in figure 1, the ATP-PC system is extremely important for exercise lasting less than 10 seconds and anaerobic glycolysis is extremely important during high intensity exercises lasting 30-90 seconds. Students should realize, however, that exercise lasting longer than a few seconds requires a significant amount of aerobic ATP production (see figure 1 and appendix page 77). For example, when a physiologist refers to "anaerobic exercise" they mean that most of the energy needed to perform the exercise comes from anaerobic sources; it does not mean that all of the energy comes from anaerobic energy sources. Glycogen & Glucose Glycolysis NADH + H+ Pyruvate NADH + H+ & FADH2 NADH + H+ & FADH2 Fatty acids Beta Oxidation Acetyl Co A Citric Acid Cycle NADH + H+ & FADH2 from all of these processes, along with oxygen, are used to form ATP via the Electron Transport Chain Mitochondria Figure 3. Highly simplified figure depicting the aerobic production of ATP from fats and carbohydrates. Whereas fats can not be used anaerobically to produce ATP, aerobic energy systems can use both fats and carbohydrates as substrates to produce ATP. The mitochondria are where the important aerobic metabolic processes(-oxidation, the citric acid cycle, and the electron transport chain) take place. In general fat metabolism is increased by: exercise training, increasing the duration of the exercise, decreasing the intensity of the exercise, or eating a diet rich in fats. On the other hand, high intensity exercise, inactivity, carbohydrate ingestion, and heat stress tend to increase reliance on carbohydrates. Fatty acids are broken down to form acetyl CoA by a process called -oxidation (betaoxidation). The acetyl groups from Acetyl CoA, formed either from glycolysis or from oxidation, can then enter the citric acid cycle (also called the tricarboxylic acid cycle or the Krebs cycle). The citric acid cycle, glycolysis, and -oxidation all result in the formation of reducing Lab IV - 5 equivalents (either NADH + H+ or FADH2, or both). The electron transport chain uses the electrons and hydrogen ions from these reducing equivalents to create a large concentration gradient for H+ across the inner mitochondrial membrane. The energy created by this large concentration gradient can then be harnessed to drive the formation of ATP. Oxygen serves as the final electron acceptor in the electron transport chain such that oxygen, the electrons, and the hydrogen ions ultimately form water (H2O) in the process of making ATP. This aerobic production of ATP in the mitochondria is referred to as oxidative phosphorylation. It should also be noted that one ATP is also indirectly produced during each round of the citric acid cycle. It is beyond the scope of this class to cover the details of glycolysis, beta oxidation, the citric acid cycle, and the electron transport chain. However, students are encouraged to review these processes, which were covered in their introductory biology, human physiology, and exercise physiology lecture courses. Is it a disadvantage that the aerobic energy systems, which can produce ATP for prolonged periods of time can not produce ATP at high rates, or that anaerobic energy systems can produce ATP very fast but not for very long? No, because there is no single energy system that could allow for both very high rates of ATP production and long-term sustainable ATP production. It is the presence of these multiple energy systems that allows humans to perform exercise at such varying intensities and durations. The contrasting properties of our energy systems allows us to exercise at very high intensities when necessary and maintain moderately high rates of energy expenditure for prolonged periods of time when necessary. These energy systems and metabolic pathways have evolved over a long time (unicellular eukaryote fossils have been traced back to 1.8 billion years ago!). In our more recent evolutionary past (10,000 years ago), it would have been necessary for our hunter-gatherer ancestors to be able to move both very quickly (to evade predators or capture prey) and be able to travel long distances (for gathering food stuffs). Thus, it is no accident that humans have retained energy systems for both quick and for prolonged physical activity. In summary, while the anaerobic energy systems can produce ATP much faster than the aerobic systems, they can not produce ATP at these very high rates for very long. Thus, when high power outputs are required, the anaerobic systems are absolutely necessary in order to meet the cell's ATP demands (although aerobic energy systems contribute significantly to any exercise lasting more than a few seconds). On the other hand, while aerobic energy systems can not produce ATP as fast, they can produce ATP at moderately high rates for long periods of time and can use both fats and carbohydrates. Intracellular Regulation of Energy Metabolism During Exercise The cell's metabolic pathways are regulated by multiple complex mechanisms, including both intracellular and endocrine regulation. Dozens of enzymes catalyze the important steps involved in the cell's metabolic pathways. The most important sites of energy system regulation are at the rate limiting steps of these energy systems. A rate limiting step is the slowest step in a series of reactions. For example, consider a series of reaction in which A is converted to D in a series of steps: ABCD If the rate limiting step in the following series of reactions is the formation of C from B, then speeding up, or slowing down, the transition of A to B or C to D will have no effect on the overall process. The only reaction that has the potential to speed up or slow down the conversion of A to D is the rate limiting step (B to C). The enzymes that catalyze the rate limiting steps of metabolic pathways are therefore referred to as rate limiting enzymes. For example, the rate limiting enzyme for glycolysis is phosphofructokinase (PFK). If PFK is stimulated then glycolysis proceeds more quickly, if PFK is inhibited then glycolysis proceeds more slowly. Fortunately for students, the intracellular factors that help to regulate cellular metabolic pathways generally make sense. For example, an increase in ATP tends to inhibit the rate limiting enzymes of glycolysis (PFK), glycogenolysis (phosphorylase), and the citric acid cycle (citrate Lab IV - 6 synthase and isocitrate dehydrogenase). On the other hand, an increase in ADP or AMP tends to stimulate glycolysis, glycogenolysis, the citric acid cycle, as well as the electron transport chain. This makes sense; if there is a sufficient quantity of ATP available to meet the cell's needs, why make more? On the other hand, an increase in ADP or AMP suggests that the cell's energy demands are not being met, and therefore the processes that make ATP need to speed up. Thus, the ATP/ADP ratio is a very potent regulator of the cell's energy producing processes. Calcium and hydrogen ions also play important roles in intracellular regulation of the metabolic pathways. The increase in cytoplasmic calcium during muscle contraction is also an important stimulus for some of these metabolic pathways. For example, calcium stimulates both glycogenolysis (by stimulating phosphorylase) and the citric acid cycle (by stimulating isocitrate dehydrogenase). A drop in pH (an increase in H+) inhibits PFK activity and thus slows down glycolysis. This means that during high intensity exercise, if lactate and H+ start to accumulate, glycolysis will likely slow down. The metabolic pathways also interact with each other. For example, when a lot of fatty acids are being broken down (e.g. during low intensity exercise), it results in large quantities of acetyl CoA, which speeds up the citric acid cycle, and increases cellular concentrations of citrate (the first product produced in the citric acid cycle). High citrate concentrations can slow down glycolysis and reduce the use of carbohydrates. On the other hand, when glycolysis is proceeding very quickly (e.g. during high intensity exercise) H+ concentrations in the body increase and inhibit lipolysis (by inhibiting hormone sensitive lipase), thus reducing fat utilization as a fuel source. These types of interactions are partly responsible for the fact that at low intensities we tend to rely more on fats as a fuel source and less on carbohydrates as a fuel source. Endocrine Regulation of Energy Metabolism During Exercise The endocrine system is also extremely important in regulating our metabolic pathways (see table on appendix page 78). Insulin, which is secreted from the -cells of the pancreas, 1) stimulates uptake of fuel sources (glucose, fatty acids, amino acids), 2) promotes storage of these fuel sources (glycogenesis, lipogenesis, and protein synthesis), and 3) tends to reduce the breakdown of glycogen and proteins. Glucagon (secreted by -cells of the pancreas), epinephrine and cortisol (secreted by the adrenal gland), and growth hormone (GH, secreted by the pituitary gland) have slightly different actions, but tend to promote mobilization of glucose and fatty acids (by stimulating glycogenolysis and lipolysis); increasing the delivery of these important fuel sources to the active skeletal muscle. During exercise insulin secretion decreases and the secretion of glucagon, epinephrine, cortisol, and growth hormone all increase. The combination of these changes in hormone concentrations results in 1) an increased breakdown of fuel stores (e.g. glycogen and triglycerides) and 2) a mobilization of fuel substrates (e.g. glucose and fatty acids) that can be used by the active tissues. When comparing trained and untrained individuals, trained individuals will usually have less of a decrease in insulin and less of an increase in glucagon, epinephrine, cortisol, and GH at any given exercise intensity. As a result, glycogen will not be broken down as quickly in trained individuals and they will tend to rely more on fats than untrained individuals. High exercise intensities result in a large increase in circulating epinephrine and a large decrease in insulin. This results in a large increase in glycogen breakdown, and therefore increases carbohydrate metabolism and decreases fat metabolism. As you can see, changes in circulating hormone concentrations are partly responsible for the changes in fat and carbohydrate metabolism that occur with exercise training or with varying exercise intensities. Thermoregulation and Exercise The human body and the physiological processes within it function best when the temperature is maintained at around 37C. A significant increase or decrease in body temperature can be fatal. The hypothalamus is responsible for regulation of body temperature. Heat is Lab IV - 7 generated by the metabolic processes in the body, and is dissipated at the skin. At rest, most of the body's heat is given off by radiation, whereas during exercise most of the body's heat is given off by evaporation. The body's core temperature is usually around 37C, but it is not uncommon for this to increase somewhat during exercise. In order to dissipate heat from the core to the skin, the skin temperature must be lower than the core temperature. That is, there must be a thermal gradient between the core and the skin in order to transfer heat from the core to the skin. The body's thermal gradient can be calculated as core temperature minus skin temperature. During exercise in a normal environment (not too hot or too humid) the skin temperature decreases, which increases the thermal gradient and prevents core temperature from increasing too much. However, if the subject is exercising in a hot humid environment (or wearing too much clothing), evaporative cooling of the skin will not be as effective, and the body will have a hard time dissipating heat. If the body can not dissipate heat, skin temperature increases, decreasing the thermal gradient and reducing the transfer of heat from the core to the skin. As a result, core temperature will now start to increase more rapidly; increasing the danger of developing heat-related illnesses. It also should be noted that in a hot, humid environment, more blood must be sent to the skin in order to transfer the heat from the core to the skin; thus heart rate and cardiac output will need to be higher at any given exercise intensity in a hot humid environment. Heat also effects energy metabolism. Heat stress tends to increase the use of carbohydrates and by accelerating glycogen breakdown and speeding up anaerobic glycolysis. LABORATORY OBJECTIVES: 1. To help students better understand how exercise training, intensity, duration, thermal stress, and carbohydrate ingestion influence exercise metabolism: 2. To help students understand how exercise training, intensity, duration, thermal stress, and carbohydrate ingestion affect the relative contributions of the major metabolic pathways to exercise energy expenditure. 3. To help students understand how exercise training, intensity, duration, thermal stress, and carbohydrate ingestion influence the active muscles' use of fats and carbohydrates as fuel substrates during exercise. 4. To help students understand the body's thermoregulatory systems and the physiological consequences of thermal stress. 5. To be able to calculate the amount of energy coming from fats and carbohydrates during different exercise bouts LABORATORY PROCEDURES & CALCULATIONS Assess VO2, RER, blood glucose, and blood lactate during the following. Using the RER, determine what percent of kcal are coming from fat and what percent are coming from carbohydrates. Calculate the total number of kcal coming from fat and from carbohydrates. Each group will be assigned one or more of the following experiments: Experiment 1. Effects of Intensity Same subject: Two 10min bouts of exercise at~50% and ~75% of max. Take measurements during the last minute of exercise. Experiment 2. Effects of Duration Same subject: One continuous exercise bout at ~60-70% VO2max. Take measurements at ~5 minutes into the exercise bout and after 30-35minutes of exercise. Experiment 3. Effects of Training: Use two subjects of the same gender and who are approximately the same size. Use one subject who does not run (or cycle) and one subject who does run (or cycle). Both subjects will exercise at the same absolute exercise intensity (recommend ~90-120 Watts if subjects are female or ~120-150 Watts if male) Lab IV - 8 Experiment 4. Effects of Thermal Stress: Same subject: Two 10 min bouts of exercise at ~75% of max. The first bout should be performed while wearing shorts and a T-shirt. The subject should put on multiple layers of clothing for the next exercise bout. Take measurements during the last minute of exercise. Additional data to be collected include heart rate, skin temperature, core temperature, and thermal gradient. Experiment 5. Effects of Carbohydrate ingestion: Same subject: Use a subject who has not eaten for at least 3-4 hours, preferably longer. Two 10 minute bouts of exercise, performed at a moderate intensity (approximately 60-70% of maximum) before and then one hour after ingesting a drink containing carbohydrate. 1. Collect your subject's expired air during rest for 5 minutes and collect the necessary data to calculate resting oxygen consumption in L/min. 2. Collect expired air during the final minute of exercise (except for the first collection in experiment 2, which should be taken at 5 minutes) to insure steady state exercise. Following analysis of collected air (as above) a second exercise bout will be performed. Your instructor will assign you an ergometer and intensity. 3. Calculate resting VO2 and exercise VO2 as you have done in previous labs. Then calculate the Net VO2 for each of the exercise bouts as you did in the previous lab. VO2 = VEstpd (NF x 0.2093 - FEO2) where NF = (1-FEO2+ FECO2)/.7904 Net VO2 (L/min) = Gross VO2 - Rest VO2 (both in L/min) 4. Calculate the RER during the exercise bouts, and determine the caloric equivalent from table in the Appendix. Calculate energy expended as follows: RER = VCO2 / Gross VO2 *look at RER table for Kcal/L Net VO2 (L/min) x caloric equivalent (Kcal/L) = Energy expended/min (Kcal/min) Energy expended (kcal/min) x minutes of exercise = Energy expended (kcal) 5. Using your RER table to determine what percent of Kcal came from fats and from carbohydrates. Determine the total Kcal from fat and carbohydrates as follows (remember to convert the percent values to decimals for your calculations). You do not need to perform these two calculations for experiment # 2 (effects of duration). Total kcal from fat = Energy expended (kcal) x % of kcal from fat Total kcal from carbohydrate = Energy expended (kcal) x % of kcal from carbohydrate 6. Alert your instructor at the end of each exercise for blood lactate & glucose determination. 7. For the thermal stress experiments, experiment #4, also collect heart rate, skin temperature, core temperature, and thermal gradient information. Also collect environmental temperature and relative humidity information using the sling psychrometer. Thermal gradient can be calculated as follows: Thermal gradient = core temp – skin temp Lab IV - 9 Experiment # _______ Subject Weight(s) __ Water Vapor Pressure Kg Ambient Temp. mmHg STPD Correction Factor _______________ Rest a. FEO2 b. FECO2 c. Sample Volume (L) d. Meter Volume (L) e. f. ATPS Volume (L) VE min. Vol. ATPS (L/min) g. VE STPD (L/min) h. NF i. Gross VO2 (L/min) j. VCO2 (L/min) k. l. RER Caloric equivalent (Kcal/L) (from RER chart) m. Net VO2 (L/min) n. = Gross VO2 - rest VO2 Energy Expended/min (kcal/min) = Net VO2 x Caloric Equiv. o. p. q. °C Ambient Pressure Energy Expended (kcal) % of Kcal from fat (from RER chart) % of Kcal from carbs (from RER chart) Total Kcal from fat = E. Exp x % from fat Total Kcal from carbs = E. Exp x % from carbs Blood Lactate Blood Glucose r Lab IV - 10 Exercise 1 Exercise 2 mmHg Experiment # _______ Subject Weight(s) __ Water Vapor Pressure Kg Ambient Temp. mmHg STPD Correction Factor _______________ Rest a. FEO2 b. FECO2 c. Sample Volume (L) d. Meter Volume (L) e. f. ATPS Volume (L) VE min. Vol. ATPS (L/min) g. VE STPD (L/min) h. NF i. Gross VO2 (L/min) j. VCO2 (L/min) k. l. RER Caloric equivalent (Kcal/L) (from RER chart) m. Net VO2 (L/min) n. = Gross VO2 - rest VO2 Energy Expended/min (kcal/min) = Net VO2 x Caloric Equiv. o. p. q. °C Ambient Pressure Energy Expended (kcal) % of Kcal from fat (from RER chart) % of Kcal from carbs (from RER chart) Total Kcal from fat = E. Exp x % from fat Total Kcal from carbs = E. Exp x % from carbs Blood Lactate Blood Glucose r Lab IV - 11 Exercise 1 Exercise 2 mmHg Thermal stress – other data 1. determine % humidity using sling psychrometer wet bulb temp: _______ dry bulb temp: _______ % humidity: _______ 2. Record heart rate, skin temperature, core temperature, and thermal gradient during each minute of exercise: Time (min) 1 2 3 4 5 6 7 8 9 10 Heart rate Normal Core Skin temp temp Thermal gradient Heart rate Thermal Stress Core Skin temp temp Thermal gradient Class data % Kcal RER fats Experiment #1 - intensity %Kcal carbs Energy expended ~ 50% ~75% Experiment #2 - duration at 5 minutes at ~30 minutes Experiment #3 - training trained untrained Experiment #4 - Thermal stress normal Heat stress Experiment #5 - Carbohydrate ingestion fasted after ingestion Lab IV - 12 kcal from fats kcal from carbs Blood glucose Blood lactate In lab questions: 1. How (and why) were RER and the percent of kcal from fat and carbohydrate affected by each of the following? Exercise intensity Exercise duration Exercise training Thermal stress Carbohydrate ingestion 2. How (and why) were the total kcal from fat and carbohydrate affected by each of the following? Exercise intensity Exercise training Thermal stress Carbohydrate ingestion 3. How (and why) were blood glucose affected by each of the following? Exercise intensity Exercise duration Exercise training Thermal stress Carbohydrate ingestion Lab IV - 13 Study Questions 1. What happens to skin temperature, core temperature, and thermal gradient during normal exercise? What happened to these variables (how did they change) during exercise in a hot humid environment (such as when wearing lots of clothes)? Explain why these variables follow these trends. 2. Did heart rate stay that same between normal and thermal stress conditions? Explain any observed differences. 3. Fill in the following table regarding the major hormones that influence exercise metabolism. Hormone secreted from major metabolic actions effect of acute exercise effect of chronic exercise 4. How do the hormonal changes that occur with acute exercise (from table above) influence exercise metabolism? 5. How do the hormonal changes that occur with chronic exercise (from table above) influence exercise metabolism? 6. If a person wanted to lower their percent body fat, would you recommend a high or a low intensity, or does intensity matter? Lab IV - 14