Functions of Photosynthetic Reaction Center and Structural
advertisement

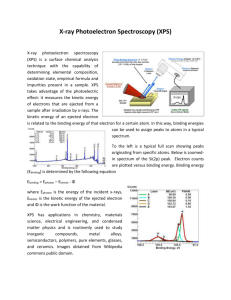
Ahmet Selim Vakkasoglu Functions of Photosynthetic Reaction Center and Structural Changes in QB Site Introduction: In photosynthesis, the important issue is the trapping of light energy. In most plants and photosynthetic bacteria chlorophyll is the photoreceptor molecule. Chlorophylls are polyenes that means they have alternating double and single bonds. This property makes them photoreceptors. They have absorption bands at visible region. This is the best region to absorb the sunlight that comes to the earth’s surface. Adsorbed light excites an electron and if this electron cannot find a suitable acceptor, then it relaxes back and emits heat. So this excited electron is given to an acceptor and a couple of positive and negative charges are produced. This process is called photoinduced charge separation. This process occurs in reaction center. Reaction center is a membrane protein which contains three subunits, L, M, and H. It has four kinds of cofactors; four bacteriochlorophylls, two bacteriopheophytins, two quinones and one non heme high spin Fe2+. (FIG1) L and M subunits are the structural and functional ones. They are positioned mostly in the transmembrane region of the protein. Each have five membrane spanning helices where H subunit has only one. H subunit lies on the cyoplasmic side of the membrane. Bacterichlorophylls (Bchl) are similar to chlorophylls with minor differences which shift absorption wavelength to near infrared. Bacteriopheophytin (BPh) has two protons instead of the magnesium ion at bacteriochlorophyll`s center. Figure 1: Bacteriochlorophyll (on the left) and bacteropheophytin (on the right) Function: Chemical reaction starts with the absorption of light by the Bchl-b dimer. This dimer is also called special pair due to its fundamental role in photosynthesis. Special pair is located near to the periplasmic side of the membrane. It absorbs at 960nm so it is often referred as P960. Absorption results in ejection of an electron. This electron is transferred to the BPh in the L subunit. This electron transfer occurs in less than 10psec. At this point positively charged P960+ and negatively charged BPh- can recombine, which conversion of the adsorbed light energy into heat. Three factors can be listed to answer the question “why do not recombination of the charges occur?”. Firstly, there is another acceptor, (QA), the tightly bound quinone. It is positioned at less than 10A away from BPh-. Secondly, the reduced heme in cytochrome is less than 10A away from the Bchl dimer. It`s known that electron transfer strongly depends on the distance. So these two factors seem to be challenging. The third one is the rate of the electron transfer reaction from BPh- to P960+. The rate of this reaction is slower than the reverse reaction. Thus, electron proceeds to QA from BPh-. After the electron reaches to QA, it proceeds to QB, loosely bound quinone. Repetition of this electron transfer beginning with the absorption of a photon results in the reduction of QB from Q, quinone, to QH2, quinole. These protons are taken from the cytoplasm and cause the development of a proton gradient across the cell membrane. In this report, the function-structure relationship of loosely bound quinone, QB, in charge neutral (DQAQB) and charge separated (D+QAQB-) states will be discussed. (where D is the primary donor, dimer). In addition to this, the possible pathways of proton uptake from cytosol will be shown. In the rest of the report QB binding site will be discussed, because the other cofactors like, Bchl, BPh are structurally more conserved during the electron transfer. The electron transfer rate from primary ubiquinone QA- to secondary ubiquinone QB is increased when the RCs are frozen under illumination which means the charge separated state D+QAQB-, when compared to RCs frozen in dark. In Figure 2, the QB binding pocket is shown, both in dark and light structures. There are two binding structures in the dark state, QB1 and QB2. The light structure is very similar to the QB2 one. Ubiquinone is positioned in the binding pocket with O1 of QB, 6.72A away from the nitrogen of the His L190. The other carbonyl oxygen of the quinone, O4, form a H-bond Figure 2: O1 to His 190 distance is 6.72A. O4 to Ile 224 backbone distance is 3.14A with amide backbone of Ile L224. The ubiquinone ring lies parallel to Phe L216 (Figure 3). This residue is conserved and contributes to the binding affinity of the ubiquinone. Actually there are several published papers (Ermler et. al., Chang et al., Arnoux et al., Allen et al.) claiming different positions for QB binding pocket. These discrepancies can be due to the electrons created by the X-ray irradiation during data collection. These electrons can reduce the ubiquinone in the QB binding pocket. The structure of the charge separated (light) state is similar to the dark one. Here, carbonyl oxygen, O4, of the ubiquinone makes a H-bond to His L190. (Figure 4) O1 carbonyl oxygen makes another H-bond to the backbone amide nitrogen of Ile L224. In Figure 3: Phe L216 lies parallel to quinone ring. addition to these two H-bonds, O1 can have two other H-bonds to Ser L223 and to the backbone of L225, where the distances to O1 are 3.21A and 3.27A, respectively. The most important difference between light and dark structures of the ubiquinone is the movement of the center of ubiquinone by 4.5A toward the cytoplasmic region.. Because of the proton uptake is done in the light state (D+QAQB-), this movement enables quinone to come to closer to the cytoplasmic side and take the protons easier. Another change in the structure is the twist of the isoprene tail by 180o. It is known that electron transfer is achieved in the samples of RCs frozen under illumination where it is not observed in the samples of RCs which are frozen in the dark. Up to this point two positions of the QB binding pocket is told. One is the structure that the electron at QA- cannot be transferred to QB, that is, transfer is inhibited. The other Figure 4: Bond distances beginning from the upper left 2.96A, 3.21A, 3.17A, 2.81A. enables the electron transfer. In dark, QB1 is thermodynamically favored. Electron transfer pathway is cut in this structure, because the 4.5A moved position of the quinone unables it to have H-bond to His L190. The way electrons followfrom QA- to QB includes His L190. So in QB1 electron cannot jump to quinone where in QB2 it can. So movement of ubiquinone from QB1 to QB2 is a necessary step for the electron transfer from QA-QB to QAQB-. From the previous experiments it is known that the rate limiting step is not the electron transfer (Graige et al.) These experiments are done by changing the kind of ubiquinone in the QA site. Thus, the rate limiting step is the conversion step between these two states, QB1 and QB2. So energy barrier between these states becomes very important. Conversion of the ubiquinone (Q) to ubiquinol (QH2) requires 2 electrons. So a second electron is transferred in the same way and reduces the ubiquinone. The formed ubiquinol leaves RC, goes to quinol pool in the membrane. So the protons taken from cytoplasm form a proton gradient. At this point, the way that the protons follow from cytoplasm to the quinone binding site is important. Normally it is accepted that the protons follow proton donor and accepter groups. These groups can be the side chains of aminoacids and the water molecules. Although several aminoacids that are used in proton uptake are known by site directed mutagenesis, the positions of the water molecules are not well known. They require higher resolution. Two possible proton pathways are indicated in Figure 5 as P1 and P2. Pathway P1 lies along ~23A from QB binding pocket to the cytoplasmic side. This pathway is perpendicular to the membrane surface. It includes Ser L223, Thr L226, Glu L212, Asp L210, Lys H130, Glu H122, Glu M236, His H68 and Arg H70 (Figure 6). The second pathway, P2, follows the interface between H and M subunits parallel to the membrane surface. In this pathway, Ser L223, Glu L212, Asp L 213, Glu H173, Asn M44, Ser M8, Gln H174 and Gln M11 exist (Figure 7). End of this pathway is at the negative charged membrane where proton concentration is higher than bulk water. Figure 5: Proton pathways P1 and P2 beginning from QB binding site. Next and the final step is the release of the quinol and replacement by ubiquinone. For this replacement the binding affinities of three species QBH2, QB- and QB are important. Firstly binding affinity of the QB- should be higher than others. Because release of this chemically reactive ubisemiquinone would be detrimental. It has to be kept due to its functionality. Binding affinity of QB is expected to be higher than the QBH2 for the replacement. Experiments showed that this is true (Wraight et al.). So the binding affinities are in the decreasing order of QB- > QB > QBH2. These results are in accordance with the structures shown here. As it is told above, QB- has 3 H-bonds by both carbonyl oxygens, where the QB has only one carbonyl oxygen having H-bond. In the case of the ubiquinol, both carbonyl oxygens are protonated, resulting in weaker binding. Figure 6: P1 proton pathway goes perpendicular to the membrane surface. Figure 7: P2 proton pathway lies on the interface of H and M subunits. Reference: 1) M. H. B. Stowell, T. M. McPhillips, D.C. Rees, S. M. Soltis, E. Abresch, G. Feher, Science, 276, 1997, p812. 2) E. Takahashi, C. A. Wraight, Biochim. Biophys. Acta, 1020, 1990,107 3) M.S. Graige, G. Feher, M. Y. Okamura, Biophys J. 70, 1996, Abstr 10 4) “ Structure and Function of the Reaction Center From Rhodopseudomonas Sphareoides” G. Feher, M. Y. Okamura. 5) Biochemistry, J. Berg, J. Tymoczko, L. Stryer, 5th edt. Freeman, 2001 6) “The Photosynthetic Reaction Center”, J. Deisenhofer, J. R. Norris, Vol I & II. Academic Press, 1993.