Primer Part II
advertisement

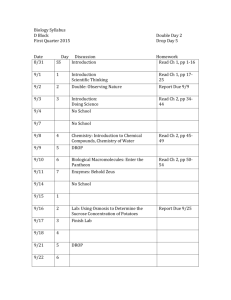
Almost all errors in analysis occur at this stage of converting the model into a matrix. The following matrix formally identifies each element. a1 1 a1 2 a1 3 a2 1 a2 2 a2 3 a 3 1 a3 2 a3 3 MATHCAD procedures If you have access to MATHCAD, a model is entered and analyzed in the following way. Using a simple tiered food-chain model, say of a self-regulated plant 1, its herbivore 2 and a specialized predator 3, hand draw the conceptual model. This type of representation is called a signed digraph (for di-rected graph). From this graph, the community matrix can be represented symbolically. Locate the Matrix icon (View Toolbars Matrix or Ctrl-M) and call up a 3x3 matrix. Into each element (appearing as black squares), enter the following, corresponding to the diagram above (do not use subscripts for this analysis). Here -a11 (i.e. negative link to 1 from one) represents the a11 a12 0 a21 0 a23 0 a32 0 self-effect on variable 1, -a12 is the negative link to 1 from 2, a21 is the positive link to 2 from 1, and so on. This matrix represents the set of Lotka-Voltera simultaneous equations for this system. Practice the following operations. On each diagonal element, add ‘-’ by entering –L (in lower case), selecting L and pressing Ctrl-G (changes font to Greek alphabet). The solutions to the symbol are called eigenvalues. Eigenvalues are analogous to the growth rate and represent the recovery characteristic of each population following a disturbance. a11 a12 0 0 a23 a21 0 a32 0 Then ‘grab’ the whole matrix, through either dragging the pointer or pressing the space bar if your cursor is already in the matrix. Calculate the determinant from Symbolic Matrix Determinant. You will get a complicated looking expression. 2 3 a11 a11 a23 a32 a23 a32 a21 a12 Place your cursor on the key in Symbolic Collect and the polynomial form will be obvious; this result is the characteristic polynomial, (analogous to the Euler equation) and solutions to are the eigenvalues. 3 2 a11 (a21 a12 a23 a32) a11 a23 a32 Keeping your pointer on the , key in Symbolic Polynomial Coefficients. The resultant column vector lists the coefficients of the characteristic polynomial, which are the values a11 a23 a32 a21 a12 a23 a32 a11 1 1 of feedback at various levels, from F0 = -1 upwards to F4. For even numbered systems, of 4 variables for example, multiply the vector by –1 for the correct result (select the vector, then key in *-1; select the whole expression again and Shift-F9). Feedback will be the basis for assessing the presence of a stable equilibrium. As a preliminary evaluation, all feedback must be negative for stability to occur, which is the case here. The limit of matrices in MATHCAD is 10x10,although it may not be able to solve symbolically systems much smaller. The workshop will introduce MAPLE techniques to handle very large system. Practical model building advice Mathematical modeling is considered essential in modern ecology and conservation biology. A mathematical model should be conceived as a hypothesis, not perceived a result. The difference from a more general conception of a hypothesis is that a mathematical construct gives a narrow and therefore very rigorous hypothesis. Statistics is one form of mathematical modeling that all scientists have come to accept, but relies heavily on data and does not deal with large systems very well. Qualitative modeling is a formal technique that draws inferences from community structure and is well suited for the types of problems that face community ecologists. Some rules of thumb, the so-called Quirk-Ruppert rules, to keep in mind when analyzing or building models are: -at least one diagonal entry ( a11…) must be negative, or self-regulated, for stability to occur -a system with a row or column of zeros is unstable -interference or mutualism between two species reduces stability due to their positive feedback. The most stable pairwise relationship is predator-prey because of its negative feedback -loops or cycles of three or more parameters introduce instability. In other words, links that skip a level, such as omnivory, introduce instability -the presence of unstable relationships does not mean that the system is unstable overall, but rather that conditions exist that must be satisfied for stability to occur Also keep in mind that: -a subsystem with overall negative feedback that is linked to a variable may be represented as a self-effect. One need not represent the whole subsystem. Thus, a predator that relies on prey outside the specific system under study would have a selfeffect attached to it. -a straight chain of predator-prey relationships, since it meets stability criteria, may be condensed into a single parameter within a larger system -a system with all interactions as negative or zero, or all positive or zero is unstable -a system with a variable linked to two or more satellite systems is unstable. A satellite is a subsytem of zero feedback linked to the system through a single variable, its principal. This is actually easy to tell because the row and column corresponding to the satellite will be all zeros except at its principal. Typically, a satellite is a specialized predator; parameter 3 in the model above is a satellite of parameter 2. -a satellite will buffer the principal so that no change in other variables will affect the principal except through the satellite and no change to the principal will affect the system except the satellite. -no parameter can be devoid of an input link -there are two major types of competition. Interference or contest competition is direct and represented as double negative links. Resource or scramble competition is indirect and arises when two or more populations consume the same resource; the resultant instability gave rise to Gause’s Law, also called Competitive Exclusion Principle. 2 THE PARADOX When ‘introduced’ three decades ago, mathematical modeling of communities challenged ecology with a serious and difficult paradox, the stability-complexity paradox, wherein complexity appears incompatible with stability. The mathematical considerations that coerce this conclusion are often referred to as the Routh-Hurwitz criteria. The intuitive and so-called Eltonian perspective that had predominated until then had asserted that stability of ecosystems is maintained because of complexity. Partly as a result of numerous attempts to circumvent this paradox, stability has become a vague term in ecology and 100+ definitions have been suggested (Grimm and Wiesel 1997). The community matrix approach is powerful because it reduces the approach to Malthusian parameters and therefore is interpretable from an evolutionary point of view. It also sets formal definitions as a starting point. We shall discuss over the workshop how this approach can be extended to deal with the paradox. 3 EXAMPLES R 1 Ex. 1: Plankton model (from Stone 1990). Plankton (P) and bacteria (B) consumes a common resource (N: nitrogen), while plankton leach carbon to bacteria. Two predators, rotifers (R) and zooplankton (Z), prey on them and leach nitrogen back into the system when they die. Z 3 B 2 P 4 N 5 M 1 Ex. 2: Old field model (Schmitz 1990). Four plants (F, P, S, H) compete for nitrogen (N) and interfere with each other as well. A grasshopper (M) consumes each plant. Schmitz completely specified quantities of the community matrix and then pressed the system. F 2 P 3 S 4 H 5 N 6 Ex. 3. Danish shallow lakes (Jeppesen, 1998). A complex food web of five trophic levels, ranging from nutrient, macroplankton, phytoplankton, invertebrates, zooplankton, cyprinids, predatory fishes and various aquatic birds. Eutrophication is considered a bottom-up press on nutrients Ex. 4. Snowshoe hare field press (Krebs et al. 1995, model from Dambacher et al. 1998). Plants (1) are consumed by hares (2) who are in turn preyed upon by various predators (3). The predators have other important sources of prey and are therefore ‘selfregulated’. Krebs pressed the system in various ways. Dambacher suggested a bottom-up commensal link between plants and predators, representing the likely benefit that plant cover provides to predators. Ex. 5. Scallop harvest regulation model (Puccia and Levins 1985). In order to preserve Nantucket scallops, two models are proposed. In one, the commission responds to declining population by enhancing juvenile breeding. In the other, it reduces harvest pressure by increasing the cost of permits. Which will be effective? 4 SELECTED REFERENCES Dambacher, J. M., H. W. Li, J. O. Wolff and P. A. Rossignol. 1999. A parsimonious interpretation of the impact of food and predation on snowshoe hares. Oikos 84: 530-532 Goodman, D. 1975. The theory of diversity-stability relationships in ecology. Quart. Rev. Biol. 50: 237-266 Grimm, V. & C. Wiesel. 1997. Babel, or the ecological stability discussions: an inventory and analysis of terminology and a guide for avoiding confusion. Oecologia 109: 323-334 Jeppesen, E. 1998. The ecology of shallow lakes - trophic interactions in the pelagial. National Environmental Research Institute, Technical Report No. 247. Silkeborg, Denmark. Krebs, C. J., S. Boutin, R. Boonstra, A. R. E. Sinclair ad J.N.M. Smith. 1995. Impact of food and predation on the snowshoe hare cycle. Science 269: 1112-1115 Levins, R. & B. B. Schultz. 1996. Effect of density dependence, feedback and environmental sensitivity on correlation among predators, prey and plant resources: models and practical implications. J. Anim. Ecol. 65: 802-812 Levins, R. 1966. The strategy of model building in population biology. Am. Sci. 54: 421-431 Levins, R. 1993. A response to Orzack and Sober: Formal analysis and the fluidity of science. Q. Rev. Biol. 68: 547-555 Li, H. W. & P. B. Moyle. 1981. Ecological analysis of species introduction into aquatic systems. Tr. Am. Fish. Soc. 110: 772-782 May, R. M. 1973. Qualitative stability in model ecosystems. Ecology 54: 638-641 May, R. M. 1974. Stability and Complexity in Model Ecosystems. Princeton Univ. Press. 265 pp. Orzack & Sober. 1993. A critical assessment of Levins' The Strategy of Model Building in Population Biology. Q. Rev. Biol. 68: 533-546 Pimm, S. L. & J. H. Lawton. 1978. On feeding on more than one trophic level. Nature 275: 542-44 Puccia, C. J. & R. Levins. 1985. Qualitative Modeling of Complex Systems. An Introduction to Loop Analysis and Time Averaging. Harvard Univ. Press. 259 pp. Puccia, C. J. & R. Levins. 1991. Qualitative Modeling in Ecology: Loop Analysis, Signed Digraphs, and Time Averaging. Chap. 6, pp. 119-143 In, Qualitative Simulation Modeling and Analysis, Fishwick, P. A. and P. A. Luker (eds.). Springer-Verlag. NY. Schmitz, O. J. 1997. Press perturbations and the predictability of ecological interactions in a food web. Ecology 78: 55-69 Schoener T. W. 1989. Food webs from the small to the large. Ecology 70: 1559-1589 Stone, L. 1990. Phytoplankton-bacteria-protozoa interactions: a qualitative model portraying indirect effects. Marine Ecology Progress Series 64:137-145 Vandermeer, J. 1981. Elementary Mathematical Ecology. John Wiley and Sons. 294 pp. 5