ch25
advertisement

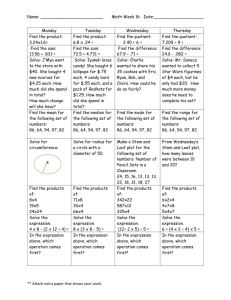
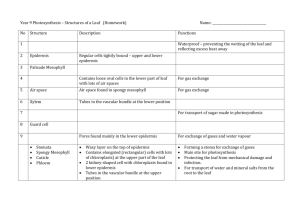
Chapter 26 THE SHOOT: PRIMARY STRUCTURE AND DEVELOPMENT FUNCTION OF THE STEM Principal functions are: Conduction. Support. Some stems are modified to perform other functions. ORIGIN AND GROWTH OF PRIMARY TISSUES. Activity of the apical meristem results in the repetitive production of phytomeres. Phytomeres consist of leaf primordia that develop into leaves and bud primordia that develop into lateral shoots. The shoot apex includes the apical meristem and the subapical region bearing the young leaf primordial. The vegetative shoot has two regions: the tunica and the corpus. The tunica forms the outermost layer of cells. These cells divide anticlinally (the newly formed cell wall is perpendicular to the surface of the stem). These divisions contribute to surface growth without increasing the number of cell layers in the meristem. The number of tunica layers varies. Most angiosperms have two tunica layers and corpus layer. These layers are designated L1, L2 and L3. The corpus consists of a group of cells that lies underneath the tunica. These cells divide periclinally (the newly formed cell wall is parallel to the surface of the stem). These divisions add bulk to the developing shoot. The bulk of the corpus cells have many vacuoles forming the central mother cell zone. The vacuolated cells are surrounded by the peripheral meristem. Anticlinal divisions: the newly formed cell wall is perpendicular to the surface of the tissue Periclinal divisions: the newly formed cell wall is parallel to the surface of the tissue http://www.vcbio.science.ru.nl/en/virtuallessons/stemgrowth/primmeristem/ Angiosperms have apices consisting of three superimposed layers of cells: two tunica layers and the initial layer of the corpus. These layers are designated L1, L2 and L3 being the innermost. L1 divides almost exclusively anticlinally but occasionally it divides periclinally. When this occurs, the inner daughter cell is displaced into the L2 layer where it differentiates as though it had been derived from the L2 layer. Similar displacements may occurs in L2 and L3 layers. The fate of a cell in the shoot meristem is dependent on its position and not on cell lineage. The corpus and the portion of the tunica overlying the corpus form the central zone, which corresponds to the promeristem of the shoot apical meristem. The central zone is surrounded by the peripheral zone, or peripheral meristem, which originates partly from the tunica, L1 and L2, and partly from the corpus, L3. The peripheral zone forms a ring around the central zone. To the inside of the ring formed by the peripheral meristem and under the central zone is the pith meristem. The cells of the central mother cell zone divide infrequently. The peripheral zone is mitotically very active. The protoderm always originates from the L1 layer. The peripheral meristem gives rise to the procambium and part of the ground meristem. The pith and the rest of the ground tissue originate in pith meristem. The shoot gives rise to leaf primordial in such a rapid succession that nodes and internodes cannot at first be distinguished. The elongation of the stem occurs by internodal elongation, which may occur simultaneously over several internodes. In some species, it occurs as a wave progressing from the base of the internode upward, while in others, such as grasses, it is restricted largely to the base of the internode. A meristematic region between two differentiated regions is called an intercalary meristem. Increase in stem thickness is the result of periclinal division and cell enlargement. In plants with secondary growth this primary thickening is moderate. In monocots, like palms, there is a massive primary growth. Growth occurs close to the apex in the region just below the young leaf bases. Localized cell division result in the formation of procambial strands. The zone of procambium formation is called the meristematic cap. The shoot apex produces the protoderm, ground meristem and procambium. PRIMARY STRUCTURE OF THE STEM There are three basic stem patterns: 1. Woody eudicots and magnoliids form a continuous ring around the pith, e.g. Tilia, basswood. 2. Herbaceous eudicots form a ring of discrete vascular bundles around the pith, e.g. Sambucus canadensis, elderberry; Ranunculus, buttercup; Medicago sativa, alfalfa. 3. Monocots have many vascular bundles scattered throughout the ground tissue, e.g. Zea, corn. Woody eudicots and magnoliids. The vascular cylinder consists of vascular bundles that are separated by inconspicuous regions of interfascicular parenchyma that connects the cortex and the pith. The epidermis is a single layer with cuticle. The stem epidermis contains fewer stomata than the leaf epidermis. The cortex contains collenchyma below the epidermis, and parenchyma cells that may contain chloroplasts. The collenchyma cells provide support for the young stem, forming a continuous cylinder below the epidermis. Primary phloem develops from the outer layers of the procambium and primary xylem from the inner ones. A single layer of procambial cells located between the primary phloem and the primary xylem never differentiates and becomes the vascular cambium. Fibers develop in the primary phloem after the internodal elongation is completed. Pith is made of parenchyma cells, some of which may have deeply colored contents. Ducts containing mucilage, a carbohydrate, are located in the pith and in the cortex. Both pith and cortex cells have large intercellular spaces, which are essential for gas exchange with the atmosphere Herbaceous eudicots. In Sambucus, elderberry, the interfascicular parenchyma, also known as pith rays, is wide separating the vascular tissue into discrete bundles around the pith. The wood is soft. The epidermis, cortex and pith are similar to those of Tilia. Protoxylem differentiates close to the pith and protophloem differentiates next to the cortex. Differentiation proceeds towards the procambium. Protoxylem and protophloem are usually destroyed during the elongation of the internode. Fibers develop after internodal elongation is completed. The stems of some herbaceous dicots have little or no secondary growth. Medicago (alfalfa) and Ranunculus (buttercup) are herbaceous. There is some secondary growth in Medicago and the structure of the stem is similar to that of Sambucus. The vascular bundles are separated by wide interfascicular parenchyma. The vascular cambium is partly fascicular and partly interfascicular. The interfascicular cambium produces sclerenchyma cells on the xylem side during secondary growth. In Ranunculus, the vascular bundles resemble those of monocots. The vascular bundles retain no procambium after the primary vascular tissue matures. Bundles that have lost their potential for further growth like in Ranunculus and monocots are said to be closed. A sheath of sclerenchyma cells usually surrounds closed bundles. Monocots. In monocots, vascular bundles are scattered throughout the ground tissue. The protoxylem and protophloem are stretched and destroyed during internodal elongation and leave a large space called the protoxylem lacuna on the xylem side of the bundle. The bundle contains two large metaxylem vessels and is surrounded by a sclerenchyma sheath. VASCULAR BUNDLES OF THE STEM AND THE LEAF Procambial strands of the stem arise behind the apical meristem just below the leaf primordia. As the leaf primordia grow in length, the procambial strands differentiate within them. From the beginning, the procambial strands of the leaf are continuous with those of the stem. The strand of vascular tissue that goes into the leaf is called leaf trace. The regions of ground tissue in the vascular cylinder above the diversion of the leaf trace is called a leaf trace gap. A leaf trace extends from its connection with a bundle in the stem called the stem bundle to the level at which it enters the leaf. A stem bundle and its associated leaf traces are called a sympodium. Buds develop in the axil of leaves and their vascular system is connected with that of the main stem by branch traces. PHYLLOTAXIS. The arrangement of leaves on the stem is called phyllotaxis. Helical or spiral. Distichous. Opposite. Decussate. Whorled. The field hypothesis of phyllotaxis. A physiological field that inhibits the growth of new primordia surrounds each primordium. New primordia will develop when growth of the stem bring the tissue outside the field of influence. The first available space hypothesis of phyllotaxis. The major factor limiting the initiation of new leaf primordia is space. As soon as new space becomes available due to distance from the summit of the previous primordia, new primordia develop. A new hypothesis suggests that biophysical forces in the growing apex determine the site of leaf initiation. The leaf primordium is initiated when a portion of the tunica surface bulges or buckles, a condition brought about in part by a localized reduction in the surface layer’s ability to resist pressure from the tissues below. MORPHOLOGY OF THE LEAF Leaves normally consist of a blade or lamina and a petiole or stalk. Scale-like stipules develop at the base of the some leaf petioles. In many monocots and some eudicots, the base of the leaf is expanded into a sheath that covers the internode, e.g. grasses. Leaves may have a simple or compound blade. Compound leaves have the lamina divided into leaflets. Leaflets can be distinguished from leaves by 1. Buds are found in axil of leaves but are absent in the axil of leaflets. 2. Leaves extend from the stem in various planes but leaflets lie in the same plane. 3. A branch with leaves has an apical bud; compound leaves usually end on a leaflet and never have an apical bud. STRUCTURE OF THE LEAF On the basis of water requirement plants can be classified as mesophytes, hydrophytes and xerophytes. The structure of the leaf is related to the habitat requirement for water. Epidermis. It is compact and provides strength to the leaf. Some xerophytes have a multiple epidermis made two or three layers of cells. It is covered with a cuticle in mesophytes and xerophytes. Cuticle is absent in most hydrophytes. The epidermis on both sides of the leaf lacks chloroplasts except for the guard cells of the stomata. Stomata (sing. stoma) may occur on the lower surface of the leaf or on both sides in mesophytes. Hydrophytes with floating leaves have their stomata on the upper side only, and those that are entirely submerged usually lack stomata entirely. Xerophytes have numerous stomata only on the lower side of the leaf often sunken in pits or depression of the leaf surface. These are called sunken stomata. Epidermal hairs may occur in the sunken stomata of xerophytes. Leaves of many plants have epidermal hairs on one or both sides of the leaf. Some of these hairs may be glandular and secrete resin-like substances. In magnoliids and eudicots, stomata are usually scattered over the surface. In monocots, the stomata are arranged in rows along the long axis of the leaf. Mesophyll. The mesophyll is the ground tissue of the leaf found in between the lower and upper epidermis. It is specialized for photosynthesis with large number of chloroplasts and air spaces. In mesophytes the mesophyll is commonly differentiated into palisade and spongy parenchyma. The palisade parenchyma consists of a layer or two of columnar cells located right under the upper epidermis and with their long axis perpendicular to the epidermis. There are intercellular spaces in between the palisade cells. In some leaves the palisade surface may be two to four times greater than the spongy surface. Chloroplasts are more abundant in palisade cells and most of the photosynthesis probably occurs in this layer. The spongy parenchyma is made of irregular shaped cell and is found on the lower side of the leaf above the lower epidermis. In some plants like corn all cells are more or less alike and there is no distinction between palisade and spongy parenchyma. In many xerophytes there is palisade parenchyma on both side of the leaf. Vascular bundle. The vascular bundles of the leaf, veins, are continuous with the vascular bundle of the stem. Magnoliids and eudicots have a netted venation. Most monocots have a parallel venation with veins extending along the long axis of the leaf. Small veins may connect the larger parallel veins in monocots. Veins contain xylem and phloem, which is primary in origin in most cases. The coarser midvein and some secondary veins may undergo limited secondary growth in magnoliids and eudicots. Commonly the xylem occurs on the upper side of the vein and phloem on the lower side. The xylem usually extends to the end of the vein but the phloem may or may not extend to the end of the vein. Veins have different degree of thickness and the minor or thinner veins play the principal role in the collection of photosynthates. Larger veins transport what the smaller veins collected. As veins increase in size, their primary function changes from collection of photosynthates to transport of photosynthates out of the leaf. Parenchyma cells with very few chloroplasts normally surround larger veins. A compact layer of cells called the bundle sheath surrounds the smaller veins. The bundle sheath extends to the end of the veins and completely surrounds the vascular tissue so it is never exposed to the air in the intercellular spaces of the mesophyll. All substances entering and leaving the vascular tissue must pass through the bundle sheath cells. In C3 grasses a second layer of small sclerified parenchyma cells, the mestome layer, lies between the much larger bundle sheath cells and the vascular bundle. Bundle-sheath extensions connect the bundle sheath of many leaves with lower and upper epidermis. The bundle-sheath extension provides mechanical support to the vein. Collenchyma and sclerenchyma cells may be present beneath the epidermis of the vein ribs or larger veins in eudicots and magnoliids. In monocots, the veins may be bordered by fibers. Collenchyma and fiber cells may found along the leaf margins of eudicots, magnoliids and monocots. Grass leaves. The epidermis of grasses is made of several cells types. Epidermal cells are commonly narrow and elongated. Bulliform cells are large cells that occur in rows and are probably involved in the rolling up during periods of drought, and unfolding of the leaf when water is available. Guard cells are thick and narrow and associated with subsidiary cells. Leaves of C3 and C4 grasses have a consistent anatomical difference. C4 grasses C3 grasses. Bundle sheath cells concentric layers (Kranz anatomy) not concentrically arranged. Mesophyll cells concentric layers not concentric layer Bundle sheath cells large with large chloroplasts small with small chloroplasts Mestome sheath absent present Mesophyll cells 2-4 between adjacent bundles many, average 12 between bundles. Photosynthates exported fast exported at a slower rate DEVELOPMENT OF THE LEAF. Genetic mosaics analysis has provide much information about the development of the leaf. Genetic mosaics or chimeras refer to plants in which cells of different genotypes occur. The difference in the genetic composition of the meristematic cells is followed through continuous cell lineages. Many variegated plants such as crotons are chimeras having defective genes for the synthesis of chlorophyll. Clonal analysis is a type of mosaic analysis that has also provided a great deal of information about the development of the leaf. In clonal analysis, individual cells of the meristem are marked and their progeny or clones are traced into differentiated regions of the plant body. Cell lineages are followed to maturity. Genes involved in the production of anthocyanins and chlorophyll have been used to follow the cell lineage that will produce variegated leaves. Leaf primordia are initiated by groups of cells in the peripheral zone of the apical meristem: It includes cells in the three layers L1, L2 and L3. It ranges from about 5-10 cells per layer in Arabidopsis, to 50-100 cells per layer in cotton and corn. These cells are called founder cells. The founder cells divide and change orientation to produce bulges called leaf buttresses. Leaf buttresses develop into leaf primordia. Dense bands of cells form on opposite sides of the primordium and give rise to the blade. The central region of the primordium gives rise to the midrib. The L1 layer gives rise to the epidermis and the L2 and L3 layers form the internal tissues. Expansion and increase in length of the leaf takes place by intercalary growth that is by cell division and cell enlargement in the various layers. Differences in the rate of growth produce the spaces and other features characteristic of different leaves. Normally growth stops first at the tip and last at the base. Maturation of the leaf is from tip to base. The characteristic growth exhibited by the leaf is said to be determinate as compared with the indeterminate growth of the apical meristems. The procambium differentiates upward into the primordium as an extension of the leaf-trace procambium. All coarse and major veins develop upward and outward toward the leaf margin. Minor veins, however, are initiated at the tip of the leaf and develop from the tip to the base. In monocots, the lateral bands of the primordium quickly grow and encircle the apex forming a hood around it. The length increases by the addition of cells at the base, the basal intercalary meristem. Elongation of cells is restricted to a zone above the basal meristem, and maturation follows as cells are pushed upwards. The difference between the sheath and the blade does not become distinct until late in the development of the leaf when the ligule appears at the top of the sheath. SUN AND SHADE LEAVE. Leaves that grow exposed to direct sunlight are smaller and thicker than those that grow in the shade are. Several layers of palisade parenchyma develop in sun leaves, with more extensive vascular tissue thicker wall epidermis. The ratio of internal surface area of the mesophyll to the area of the leaf blade is much higher in sun leaves than is shade leaves. Great variation can be found on one plant. LEAF ABSCISSION. Chemical changes in the abscission zone at the base of the petiole precede abscission of the leaf. There are two layers in the abscission zone: the separation layer and the protective layer. The separation zone is made of small cells with thin cell wall that make this layer structurally weak. Before abscission, reusable ions, amino acids and carbohydrates are returned to the stem. Enzymes break down the cell wall in the separation layer including hydrolysis of cellulose and materials in the middle lamella. Cell division may occur in the separation zone prior to abscission. If so, these new cells are the one affected by hydrolysis. Beneath the separation layer, a layer of highly suberized cells is formed further isolating the leaf from the stem. Tyloses may form in tracheary elements. Tylose is balloon-like structures produced from ray parenchyma that tend to clog and seal wounded tissue. The separation layer forms the leaf scar after the leaf falls off. TRANSITION BETWEEN VASCULAR TISSUE OF THE SHOOT AND THE ROOT. There is different distribution of vascular tissue and ground tissue in the shoot and in the root. There is much variation in the transition region between the primary body of the root and the shoot. Vascular continuity between the root and the stem is maintained throughout the life of the plant. The transition between the root and the stem is established early in development by the differentiation of the procambium. The branching and reorientation of the primary xylem and primary phloem result in the formation of a pith in the transition zone. DEVELOPMENT OF THE FLOWER. The development of the flower or inflorescence, ends the meristematic activity of the vegetative shoot apex. Transition from a vegetative to a floral shoot involves a series of physiological and structural changes in the shoot apex that transforms a vegetative shoot apex into a reproductive shoot apex. The reproductive apex is determinate and in annual means the end of their life cycle. The transition from a vegetative to floral apex I often precede by an elongation of the internodes and the early development of lateral buds below the shoot apex. The small apex with a tunica-corpus type of organization becomes broad and domelike. Commonly, the initiation of floral parts begins with the sepals, followed by the petals, then the stamens, and finally the carpels. The orders of appearance may be modified in certain flowers but the parts always have the same relative spatial relation to one another. The floral pats may remain separate during development or they may become united within whorls, connation. REGULATORY GENES DETERMINE ORGAN IDENTITY IN THE ARABIDOPSIS FLOWER. Arabidopsis thaliana is a small herbaceous angiosperm that is being used in a variety of physiological, genetic and other studies. It is like the fruit fly of the plant kingdom. A small set of regulatory genes determines organ identity in the Arabidopsis flower. Mutations that cause the formation of an organ in the wrong place are called homeotic mutations. The study of homeotic mutations in Arabidopsis has identified three classes of gene – designated A, B, and C – that are essential to the normal development and order of appearance of the floral organs produced by the flowers. Three classes of homeotic genes have been identified in Arabidopsis thaliana. 1. Class A genes function in the first and second whorl - sepals and petals. 2. Class B genes influence the second and third whorl - petals and stamens. 3. Class C genes function in the third and fourth whorl -stamens and carpels. Mutations in Class A genes sepals are converted to carpels and petals into stamens. Mutations in Class B genes petals are converted into sepals and stamens into carpels. Mutations in Class C genes stamens are converted into petals and carpels are missing. Research is still going on in order to fully understand what was said above! STEM AND LEAF MODIFICATIONS Tendrils are wire-like and aid in support, e.g. peas. Cladophylls resemble leaves and are used in photosynthesis, e.g. Christmas cactus. Thorns are modified branches used in protection of the plant, e.g. hawthorn. Stolons are slender stems growing along the surface of the ground that root and produce a plantlet, e.g. strawberry, Boston fern. Tubers are swollen underground stems used in storage and reproduction, e.g. potato. Rhizomes are elongated horizontal underground stems bearing buds in the axils of reduced scale leaves, e.g. iris. Bulbs are small conical stems with numerous storage leaves attached to it, e.g. onion, garlic. Corms are thickened rounded storage stems covered with thin papery scales, e.g. crocus, gladiolus. Stalks and petioles of some plants become thick and edible, e.g. celery, rhubarb. Thorns are modified branches e.g. hawthorn. Spines are modified leaves and spicules, e.g. cactus, barberry. Prickles are sharp outgrowths of the cortex and epidermis, e.g. rose. Some stems and leaves are modified for water storage.