Electricity of Nerves and cells

Electricity of Nerves and cells
Biological cells regulate ion movement through their membranes and thus have an electrical aspect to their operations. The cell membrane is composed of lipids and poses a barrier to the free flow of water soluble ions.
This means it has electrical resistance.
The cell membrane is studded with all sorts of channels through which various chemical species pass. Some of these channels are passive; opening or closing, but letting the concentration gradient push the species through like a temperature gradient pushes heat. Others are active pumps, using energy from ATP to push ions against the gradient.
During normal operation, cells maintain different concentrations of ions inside and outside the cells. When this leads to an imbalance in electrical charge, the membrane has an electric potential difference across it and is acting as a capacitor.
Getting a feel for the numbers.
For a nerve cell, the thickness of the cell membrane is about 7 nm and the potential difference across it is around 70 mV in the resting state.
Electric field: The electric field is calculated as V/d.
For the nerve cell membrane, this gives
E = 70 mV/7nm = 10 7 V/m, directed inward.
Resistance of membrane: Using R = L/A, with =3.6x10
7 Ω m and
L=7nm. If we use a typical cell size of 0.05mm and assume it to be spherical, the area is
A = 4πr 2 = 4π(2.5x10
-5 m) 2 = 8x10 -9 m 2
This gives a resistance of
R = (3.6 x10 7 Ω m)(7 x10 -9 m)/ 8x10 -9 m 2 = 31.5 MΩ
This is an overestimate because it ignores the ion channels.
Capacitance of membrane: The dielectric constant for lipids is about
9. For the cell measurements used above, this gives a capacitance of
C = κε o
A/d = 9(8.85x10-12 C2/N m
= 89 x 10 -12 F = 89 pF
2 )( 8x10 -9 m 2 )/ 7 x10 -9 m
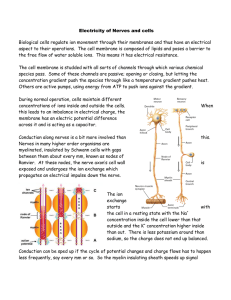
Conduction along nerves is a bit more involved than this. Nerves in many higher order organisms are myelinated, insulated by Schwann cells with gaps between them about every mm, known as nodes of Ranvier. At these nodes, the nerve axon’s cell wall is exposed and undergoes the ion exchange which propagates an electrical impulse down the nerve.
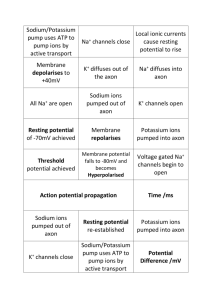
The ion exchange starts with the cell in a resting state with the Na + concentration inside the cell lower than that outside and the K + concentration higher inside than out. There is less potassium around than sodium, so the charge does not end up balanced.
The start of an impulse is the opening of the sodium channels, allowing Na + ions to flood in, changing the potential across the membrane from -70mV to
+40mV. This is followed by the closing of the sodium channels and opening of the potassium channels, dropping the potential below -70mV and then the ion concentrations are returned to the resting state by the active ion pumps.
In the nerve axon, the ions inside and outside the cell wall move along the wall from node to node, triggering the process there.
This propagation of charged ions along the cell constitutes an electric current and we can calculate a resistance for the process using the resistivity of the axoplasm inside the cell.
R = L/A, with =2Ω m and a 5 μm radius:
R for 1 cm. of axon = (2Ω m)(0.01m)/π(5x10 -6 m) 2 = 2.5 x 10 8 Ω
Notice that this is larger than the resistance of the membrane! So most charge flows out through the membrane rather than along the axon! This would mean that the recovery time for the nerve cell and therefore the speed for a pulse to propagate along it is governed by the time constant RC for the membrane which gives
RC = (31.5 MΩ)( 89 x 10 -12 F) = 3 ms
Conduction can be sped up if this process has to happen less frequently, say every mm or so. So the myelin insulating sheath speeds up signal propagation down the axon.
If we use the formulae for resistance and capacitance with the length of
1mm between nodes, we get
R = L/4πr 2 and C = 2πr L C m where C m
= capacitance per unit area = 10 -2 F/m 2
The time constant, RC is then
RC = L/4πr 2 2πr L C m
= C m
L 2 /r
And the velocity is L/RC
So, V = r/ C m
L = 10 r m/s per μm (myelinated)
= 0.1 r m/s per μm (unmyelinated)
Myelinated
2
Unmyelinated
Axoplasm resistivity (Ω m)
Capacitance/area F/m 2 membrane resistance/area Ω/m 2
5E-5
40
1E-2
0.2
Nerve Typical radius, μm
2 signal speed (m/s)
20 brain (un myelinated), type C, also some sensory nerves. muscle (motor) nerves
Type A, position, balance, delicate touch
10 (4-20) 100
Type B Sensory nerves to CNS
Giant Squid Axon, unmyelinated, L~5E-4m
3 (2-4)
1 mm = 10 3
30
100 (roughly)
Several compromises are made in the construction of nervous systems in organisms.
One is energy vs. signal speed. Larger radii nerves would give faster conduction, as seen for the giant squid neuron above, but the energy expenditure for repolarization becomes large. Increasing the spacing between nodes on a myelinated nerve increases the resistance and slows the signal, but costs less energy for repolarization.
Additionally, there is the size compromise. Only about a third of the nerves in your body are myelinated. To do so for all of them would take too much space. Your peripheral nerves would be the size of garden hoses and your spinal cord would be the diameter of a garbage can!
Sources
Martini, Fredrick H., Fundamentals of Anatomy and PhysiologyU, 7 th ed.,
Pearson, 2006, pp 400-406
Sternheim & Kane, General Physics, 2 nd ed., Wiley, 1991, Ch18
Knight, Jones & Field, College Physics, A Strategic Approach, 2007, Ch23, sections 8&9.