Removal of materials from blood
advertisement

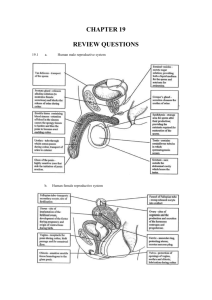
Reproduction The testes have two functions: the production of sperm in the seminiferous tubules and the production of the male hormone testosterone in the interstitial cells. The prostate gland and the seminal vesicles produce secretions which: a) b) c) d) e) provide a liquid medium (called semen) for the sperm to swim in; contain enzymes which maintain optimum viscosity (thickness) in semen; are alkaline which help to neutralise the acidic conditions in the vagina; provide nutrients for the sperm; contain hormones which stimulate contractions of the female reproductive tract. These features help to ensure successful fertilisation. Testosterone stimulates sperm production in the seminiferous tubules. It also activates the prostate gland and seminal vesicles as well as inhibiting the production of FSH and ICSH by the pituitary gland. The negative feedback effect that testosterone has on the pituitary gland means that there are relatively constant levels of testosterone, FSH and ICSH in the male. This ensures that males are continuously fertile. FSH and LH comparisons in males and females. In the testes FSH stimulates sperm production in the seminiferous tubules. In the ovaries FSH stimulates the development and maturation of a Graafian follicle and stimulates the production of oestrogen. LH is known as ICSH (interstitial cell stimulating hormone) in males. In the testes LH (ICSH) stimulates the interstitial cells to produce testosterone. In the ovaries LH brings about ovulation (release of an ovum), causes the Graafian follicle to develop into a corpus luteum and stimulates the corpus luteum to secrete progesterone. The ovaries have two functions: the production of ova and the production of the female sex hormones oestrogen and progesterone. Oviduct An ovum is surrounded by a developing Graafian follicle which, after ovulation, develops into a corpus luteum. During the first half of the menstrual cycle (day 1 - day 14) FSH stimulates the development and maturation of a Graafian follicle and stimulates the production of oestrogen in the ovary. Increasing concentrations of oestrogen promote the repair of the endometrium and stimulate the pituitary gland to secrete LH. High levels of LH cause ovulation to occur around day 14 of the cycle. During the second half of the menstrual cycle (day 14 - day 28) LH causes the Graafian follicle to develop into a corpus luteum and stimulates the corpus luteum to secrete progesterone. Increasing progesterone levels promote thickening of the endometrium and inhibit the secretion of FSH and LH by the pituitary gland. Decreasing levels of LH cause the corpus luteum to degenerate, causing levels of progesterone to decrease. By around day 28 the endometrium can no longer be maintained; it breaks down and is passed out of the body during menstruation. As the levels of progesterone decrease, its inhibitory effect on the production of FSH by the pituitary gland is reduced. The pituitary gland thus begins to increase secretions of FSH and the cycle begins again. Other changes which occur during the menstrual cycle are; a) an increase in body temperature and changes to the cervix. Body temperature can increase by about 0.5°C at ovulation. b) At ovulation the mucus secreted by the cervix becomes thin and watery as a result of high levels of oestrogen. This watery mucus allows sperm to pass through easily, thus increasing the chances of fertilisation. In females the fluctuating levels of hormones causes cyclic fertility. A woman is only fertile during the few days before and after ovulation. Intervention in fertility Some causes of infertility are: failure to ovulate; blockage of uterine tubes; failure of implantation and low sperm counts. Treatments for infertility include fertility drugs, in vitro fertilisation and artificial insemination. In vitro fertilisation occurs when ova are removed from the woman's ovaries and mixed with sperm in a laboratory. Once fertilisation has occurred, some of the developing embryos are placed inside the uterus. Artificial insemination involves injecting the male's sperm into the vagina using a syringe. Biological basis of contraception There are two biological methods of contraception: calculation of the fertile period and the contraceptive pill. Calculation of the fertile period The fertile period can be calculated by noting the rise in body temperature and noticing the cervical mucus becoming thin and watery, both of which occur at ovulation. The contraceptive pill contains synthetic progesterone which inhibits the production of FSH and LH by the pituitary gland, thus preventing ovulation from occurring. Pre- and Post-natal development Cleavage is the name given to the first few mitotic divisions after fertilisation. The cells produced decrease in size at each cleavage division. Implantation is the process during which the early embryo burrows into the wall of the endometrium and begins to form outgrowths which will form part of the placenta. Differentiation is the name given to the process by which cells begin to change into specialised cells which will form the various tissues and organs of the body. Monozygotic (or identical) twins are the result of one ovum being fertilised by one sperm. During cleavage the embryo splits and develops into two separate embryos. Since they are the product of a single zygote, monozygotic twins have identical DNA; consequently they are always the same sex. They also share a common placenta. Dizygotic (non-identical or fraternal) twins are the result of two ova being fertilised by two sperm. Such twins are no more genetically alike than any other siblings and can be the same sex or different sexes. Each twin has a separate placenta. The exchange of materials between mother and fetus occurs via the placenta. This structure consists of maternal and fetal blood vessels which are in close contact with each other but which do not allow maternal and fetal blood to mix. Useful exchanges include: oxygen and carbon dioxide which are transferred by diffusion; glucose which transferred by active transport and antibodies which pass from mother to fetus by pinocytosis. Examples of harmful substances which can pass from mother to fetus across the placenta include: pathogens like the rubella virus; drugs such as thalidomide, nicotine, alcohol and heroin. During the first three months of pregnancy the hormones oestrogen and progesterone are secreted by the ovary and corpus luteum. However about three months after fertilisation the corpus luteum degenerates and the placenta takes over the production of these hormones. Oestrogen and progesterone inhibit the menstrual cycle, in particular ovulation and menstruation. In addition they maintain the endometrium of the uterus wall and stimulate growth of the breasts in preparation for milk production. However milk production, which does not begin until after birth is stimulated by another hormone, prolactin, which is secreted by the pituitary gland. During pregnancy the placental hormones oestrogen and progesterone inhibit the secretion of prolactin by the pituitary gland. When the placenta is removed at birth the placental hormone levels suddenly drop. As a result their inhibitory effect is removed, prolactin is secreted by the pituitary gland and milk production begins. Since a fetus is not genetically identical to its mother, its cells will possess antigens which are foreign to the mother. Normally the mother's immune system will not attack the fetus because the placental barrier usually prevents any fetal cells from entering the mother's blood stream and so her immune system remains unaware of the presence of the 'foreign' fetus. Occasionally fetal cells can pass into the mother's bloodstream so that there is some mixing of fetal and maternal blood. This stimulates the mother's immune system to produce antibodies against the fetus. Although this may not damage this fetus, memory cells will persist in the mother's blood stream and, if stimulated in a subsequent pregnancy, will initiate a secondary immune response. Antibodies may cross the placenta and damage or kill the fetus. An example of this is the Rhesus factor. Oxytocin stimulates the rhythmical contractions of the muscles of the uterus wall known as labour. Birth can be artificially induced by gradually infusing oxytocin, or an artificial hormone which mimics it, into the mother's bloodstream. After birth the suckling of the new-born at the breast induces the secretion of the hormones oxytocin and prolactin by the pituitary gland of the mother. Prolactin stimulates the breasts to produce milk. It also inhibits the production of FSH and LH by the pituitary gland, thus continuing to inhibit the menstrual cycle while the baby breastfeeds. Oxytocin stimulates the ejection of milk from the glands in the breast. It also causes the uterus to contract to its normal size. Suckling and the oxytocin response are examples of a positive feedback mechanism; the more the baby suckles, the more milk is produced. Colostrum is produced for the first few days after birth. It is different from true breast milk in that it contains less lactose sugar and almost no fat. However it contains more protein, minerals and vitamins than true breast milk. Another important difference is that colostrum contains more antibodies than true breast milk. Dried cow's milk which is used for bottle-feeding also contains the nutrients needed by babies. However it does not contain any maternal antibodies, so that the baby is more susceptible to disease as it does not have the passive immunity provided by such antibodies. Sometimes colostrum and breast milk can be contaminated by substances the mother has ingested during pregnancy. Organochlorines, for example, are non-biodegradable chemicals used as pesticides on crops. An example of organochlorines is DDT. DDT molecules are fat-soluble which means they are stored in fatty tissues, including those of the breast. As milk is produced some of the pesticide passes into the milk and therefore on to breast fed babies. There are two main phases of rapid growth, known as growth spurts in humans. The first growth spurt occurs between birth and two years of age, while the second, the adolescent growth spurt, begins at puberty. Growth hormone (GH) is secreted by the pituitary gland into the blood. It stimulates the growth of bones (especially the long bones of the body in the arms and legs) and cartilage and promotes protein synthesis. An under-production of GH during adolescence leads to a reduction in growth. An over-production of GH during adolescence leads to an abnormal increase in growth, especially in the long bones. Excessive GH production in adulthood causes an abnormal increase in the bones of the hands, feet and jaw. Puberty is the name given to the processes which change the immature child into a reproductively mature adult. Many of these processes are controlled by various hormones. As well as growth hormone, the pituitary gland secretes the gonadotrophic hormones FSH and LH, which target the sex organs in males and females. As a result of increasing gonadotrophic hormones the ovaries begin to secrete oestrogen and progesterone while the testes secrete testosterone. These hormones are responsible for the development of the secondary sexual characteristics. Secondary sexual characteristics in females include: growth in height and weight; maturation of the sex organs; onset of ovulation and menstruation; increased growth of body hair; widening of hips and growth of breasts. In males secondary sexual characteristics include: growth in height and weight; maturation of the sex organs; onset of sperm production; increased growth of body hair; deepening of the voice. Transport Mechanisms All cells need a supply of nutrients and oxygen, and must get rid of waste products such as carbon dioxide. These substances pass in and out of cells by diffusion. The larger an animal, the smaller is its surface area to volume ratio. The smaller its surface area to volume ratio, the longer it takes for diffusion to occur. Because large animals like humans have a small surface area to volume ratio, diffusion is too slow a process to transport materials around the body. Therefore a transport system using vessels is required to ensure materials are transported quickly between the animal's cells and its environment. Arteries carry (normally oxygenated) from the heart to the tissues and organs of the body. They have thick, elasticated walls to withstand the pressure of the heartbeat. Arteries divide into smaller arterioles, which divide further into tiny, thinwalled capillaries. The walls of capillaries are one-cell thick, which allows the rapid exchange of materials between the blood and the tissues. Capillaries join up to form venules, which join up again to form larger veins. Veins carry (normally deoxygenated) blood back to the heart. Their walls are thinner and less elastic than those of arteries. They also contain valves to prevent the backflow of blood. Blood pressure in veins is too low to prevent blood flowing backwards. Blood pressure at the arterial end of a capillary bed is greater than blood pressure at the venous end. Thus plasma is forced out of the blood into the tissue fluid. Because red blood cells and proteins remain in the blood, the water concentration of the tissue fluid is greater than that of the blood. Water passes by osmosis back into the blood at the venous end of the capillary bed where the pressure is less. Although most of the water is returned to the blood by osmosis, about 1% does not. In order to prevent build up of this fluid in the tissues, it drains into lymphatic vessels. These eventually return the fluid to the bloodstream. Because pressure is very low in lymph vessels, they contain valves to prevent backflow of lymph. Contractions of the skeleton push lymph from one valve to the next. The heart is a muscular pump which receives blood from veins and pumps blood at high pressure into arteries. Blood flows in one direction only around the body. The heart is divided into a left and right, which are completely separate from each other. Each side is divided into an upper atrium and lower ventricle. Deoxygenated blood returns to the right atrium from the body via the venae cavae, the largest veins in the body. Blood passes into the right ventricle, from where it is pumped to the lungs via the pulmonary artery. The pulmonary artery is the only artery which carries deoxygenated blood. The blood is oxygenated in the lungs and passed back to the left atrium via the pulmonary vein. The pulmonary vein is the only one to carry oxygenated blood. The blood is passed to the left ventricle from where it is pumped out to the rest of the body via the aorta, the largest artery in the body. The atrio-ventricular valves separate the atria from the ventricles. They prevent backflow of blood into the atria when the ventricles contract. The semi-lunar valves are found at the entrance to the aorta and pulmonary artery. They prevent blood flowing back into the ventricles when they relax. The coronary artery and vein supply the heart muscle itself. The head receives blood via the carotid arteries and returns blood via the jugular veins. The kidneys major blood vessels are the renal arteries and renal veins, while the liver receives its blood from the heart via the hepatic artery and returns it via the hepatic vein. The hepatic portal vein carries blood rich in digested foods from the small intestine to the liver. Contraction of the heart muscle is known as systole, while relaxation is known as diastole. During atrial systole, the atria contract and blood passes through the atrioventricular (AV) valves into the ventricles. At this time the semi-lunar valves are closed. During ventricular systole, both ventricles contract and blood is forced out through the semi-lunar valves into the aorta and pulmonary artery. The AV valves are closed at this time. During diastole the atria and ventricles relax and the semi-lunar valves close. Blood passes into the atria from the pulmonary artery and venae cavae before the cycle begins again. Cardiac muscle is myogenic - it contracts without electrical stimulation from the nervous system. However synchronisation of the heartbeats and the rate at which the it beats is controlled by the nervous system. The heartbeat is synchronised when the atria contract together a fraction of a second before the two ventricles contract together. This synchronisation is controlled by a pacemaker and the conducting system of nervous tissue within the heart itself. The pacemaker is called the sino-atrial (SA) node and is located on the wall of the right atrium. The SA node generates rhythmical waves of nerve impulses which pass across the walls of the atria, making them contract simultaneously. The SA node also stimulates the atrio-ventricular (AV) node, which is situated between the atria and the ventricles. Nerve impulses pass down through the muscle in the septum and onto a network of fibres situated in the walls of the ventricles. This causes the ventricles to contract simultaneously from the bottom up, the most efficient way of emptying them. The SA node is attached to a part of the brain called the medulla. Nerve impulses from the medulla can cause the heart rate to speed up or slow down. The hormones adrenaline and thyroxine can also cause the heart rate to speed up. The blood pressure measured in blood vessels occurs as a result of the blood being pumped out of the heart. Blood pressure is highest in the arteries and decreases in the arterioles as a result of an increased surface area produced as a result of the greater number of arterioles. Blood pressure and the rate of blood flow decreases further in the capillaries. Blood pressure is lowest in the veins, which require valves to prevent the backflow of blood in them. Delivery of materials Haemoglobin has different affinities for oxygen under different conditions. The higher the oxygen tension the greater the affinity haemoglobin has for oxygen. The higher the temperature the lower the affinity it has. This helps to ensure that haemoglobin will release the maximum amount of oxygen to actively respiring tissues. The function of red blood cells is to carry oxygen from the lungs to the respiring cells of the body. Red blood cells are biconcave in shape, are very small , do not possess a nucleus and are flexible so that they can pass through capillaries easily. All of these features increase a red blood cell's ability to absorb oxygen and carry it to the cells of the body. Red blood cells are produced in the red bone marrow of certain bones; the sternum, ribs, vertebrae and the long bones in the arms and legs. Vitamin B12 and iron are required for the production of haemoglobin. Red blood cell have a life span of about 120 days. After this time they are broken down in the liver, spleen and bone marrow. Haemoglobin is broken down to iron and haem groups. The iron is stored in the liver for future use while the haem groups are converted in the liver to a substance called bilirubin which is excreted in the bile. The presence of villi and microvilli in the small intestine hugely increase the surface area across which digested materials are absorbed into the blood, thus maximising the efficiency of the small intestine. Carbohydrates are digested to simple sugars like glucose by amylase enzymes. Proteins are digested to amino acids by protease enzymes. Glucose and amino acids are actively transported into the epithelial cells of the villi and diffuse from here into the blood capillaries. Fats (or lipids) are digested to fatty acids and monoglycerides by the enzyme lipase. Fat digestion is promoted by the presence of bile. Bile emulsifies fats by physically breaking large fat droplets into smaller fat droplets. This increases the surface area of fat exposed to the digestive action of lipase. Fatty acids and monoglycerides diffuse into the epithelial cells of the villi where they recombine into triglycerides and exit the cells by exocytosis. The triglycerides are absorbed by the lacteals (tiny lymph vessels) in the villi and are transported to the bloodstream by the lymphatic system. Other substances which do not require digestion are also absorbed by the small intestine. Examples include vitamins and minerals like calcium and iron. Vitamin B12 can only be absorbed by the small intestine if a substance called intrinsic factor is present. The liver has a dual blood supply; the hepatic artery which supplies it with oxygenated blood while the hepatic portal vein which brings blood rich in digested foods from the small intestine. Excess glucose is converted to glycogen and stored in the liver. When blood glucose levels fall below normal glycogen in the liver is converted back to glucose. The lipid cholesterol is essential for the manufacture of steroid hormones and is a component of cell membranes. If there is insufficient cholesterol in the diet the liver manufactures it from other lipids. The liver is also responsible for the breakdown and excretion of excess cholesterol. Liver cells also store the fat soluble vitamins A, D, E and K. Excess proteins cannot be stored in the body. Instead they are broken down by a process called deamination to organic acids and ammonia. The ammonia is converted to urea transported by the blood to the kidneys and excreted by them. The organic acids can be used as a respiratory substrate by entering the Krebs cycle. The liver can convert some amino acids into others that have not been taken in in food. Glucose is transported from the liver by the blood to every living cell in the body. In the cells it is used as a source of energy which is released during respiration. Excess glucose is stored in the liver and muscles as glycogen or converted to fat and stored in fatty tissues around the body. Lipids can be used as a source of energy; fatty acids can be converted to acetyl CoA and thus enter the Krebs cycle. Lipids are also major components of cell membranes and steroids. Excess lipids are stored in fatty (adipose) tissue. In the cells amino acids are used for protein synthesis. Excess amino acids are deaminated and the urea produced is excreted by the kidneys. Minerals and vitamins have many important roles within cells including acting as enzyme activators or acting as a coenzyme. Removal of materials from blood The liver has several roles in the removal of materials from the blood, including: conservation of useful substances; detoxification of toxic materials; removal of bilirubin and its excretion as bile; production of urea. The lungs are responsible for the removal of carbon dioxide produced by all living cells during aerobic respiration. The exchange of gases (oxygen and carbon dioxide) in the lungs occurs between the alveoli and capillaries. Carbon dioxide from the blood in the capillaries diffuses into the alveoli and is exhaled. The kidneys are responsible for the removal of waste urea from the body. Urea is produced in the liver from the breakdown of excess amino acids and carried by the bloodstream to the kidneys. The renal artery carries blood into the kidney while the renal vein carries blood out. The working unit of the kidney is called the nephron. A nephron consists of a glomerulus (small knot of blood capillaries), a Bowman's capsule and a renal tubule which is surrounded by a network of capillaries. The renal tubule consists of the proximal convoluted tubule, the loop of Henle and the distal convoluted tubule. Each tubule is connected to a collecting duct which carries the urine to the ureter and from there to the urinary bladder where it is stored until it passes out of the body. Ultrafiltration occurs in the glomerulus as a result of increased pressure here. Plasma containing water, glucose, amino acids, salts and urea is forced out of the blood into the Bowman's capsule. The liquid which passes into the Bowman's capsule is called the glomerular filtrate. The high pressure in the glomerulus is produced because the blood vessel carrying blood into the glomerulus is wider than the blood vessel leaving it. Also the blood in the renal artery is itself a branch of the aorta which carries blood at a high pressure from the heart. All of the glucose, most of the salts and some of the water are reabsorbed in the proximal convoluted tubule. The glucose and salts are actively absorbed against a concentration gradient. As a result of this active transport the water concentration in the blood is lower relative to that in the glomerular filtrate. Therefore water passes by osmosis into the blood. As glomerular filtrate passes down the descending limb and round the Ubend of the loop of Henle, water continues to be reabsorbed into the blood by osmosis. As the filtrate passes up the ascending limb salt ions are actively pumped out into the surrounding tissue of the medulla. However water cannot follow by osmosis as the walls of the ascending limb are almost completely impermeable to water. In the distal convoluted tubule salts continue to be actively transported into the blood as before. At this point the cells are once again permeable to water, which passes into the blood by osmosis. As a result of the active transport of salt ions from the filtrate in the ascending limb of the loop of Henle into the tissue fluid of the medulla, the water concentration is lower in the tissue fluid than in the filtrate passing into the collecting duct. This enables large volumes of water to be absorbed from the collecting duct when necessary. The collecting ducts reabsorb enough water to restore the blood's water concentration to normal. From the collecting duct the liquid, now known as urine, passes to the ureter and then to the bladder where it is stored until passed out of the body. Regulating Mechanisms Organisms need to maintain their body within certain limits. The mechanism by which they do this is called homeostasis. Homeostatic mechanisms are regulated by negative feedback control. This produces a response to a stimulus until the stimulus has been suppressed. Blood water levels are maintained by osmoregulation. If blood water levels are low the hypothalamus stimulates the pituitary gland to secrete ADH (antidiuretic hormone). The kidneys then reabsorb water back into the blood. Small volumes of concentrated urine are produced. If blood water levels are high less ADH is secreted and less water is reabsorbed. Large volumes of dilute urine are produced. ADH makes the kidney tubules more permeable to water. If there is excess glucose in the blood then it is converted to glycogen and stored in the liver. This conversion is controlled by insulin, an endocrine hormone secreted from the pancreas. If the blood glucose levels are low the pancreas secretes glucagon which causes stored glycogen to be converted to glucose. In emergencies adrenaline is secreted from the adrenal glands and causes glycogen to be converted rapidly to glucose. This overrides the homeostatic mechanism. Diabetics are unable to produce enough insulin. The hypothalamus acts as a thermostat and regulates body temperature. It receives nerve impulses from receptors in the skin and senses temperature changes in the blood. It then sends nerve impulses to the effectors to perform the chosen response. Responses to an increase in body temperature include: vasodilation of skin arterioles; increased sweating; decreased metabolic rate; relaxation of hair erector muscles so that the hairs lie flat. Responses to a decrease in body temperature include: vasoconstriction of skin arterioles; decreased sweating; increased metabolic rate; contraction of hair erector muscles so that the hairs stand up and trap a layer of air between the skin and the outside. Voluntary responses such as putting on more clothing when cold also play an important role in temperature regulation. Hypothermia occurs when the core body temperature falls below 35°Celsius. Infants and the elderly are more at risk from hypothermia.