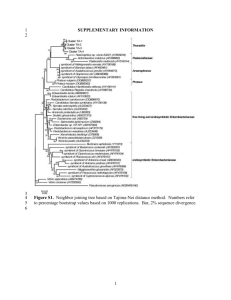
A framework for species selection: sequencing of insects to explore
advertisement

A framework for species selection: sequencing of insects to explore the evolution and maintenance of symbiotic associations Prepared by: Nicole Gerardo, Emory University (nicole.gerardo@emory.edu) Alex Wilson, University of Miami (acwilson@bio.miami.edu) Overview Organisms across many kingdoms of life are associated with microbial partners that shape their hosts' diet breadth and ability to combat disease. Some of the best insights into the establishment, evolution and maint enance of these associations have come from the study of insects associated with obligate microbial symbionts. Recently, the availability of paired insect and symbiont genomes has greatly advanced our understanding of host- symbiont interactions (Gerardo, Wilson, 2011). Here, we suggest sequencing a suite of phylogenetically diverse insects with intimate associations with microbial partners. Symbiosis is critical to defining the ecology and evolution of many arthropods (Moran, 2006). Many insect symbionts are nutritional partners, synthesizing nutrients that their hosts do not get from their diet. Thus, symbionts shape the host plant range of insects; many of these plants are agricultural crops. Other symbionts provide a role in defense against environmental stressors, parasites and pathogens. Studies of insect symbioses can therefore provide insight into multiple aspects of arthropod ecology and evolution. For example, metabo lic complementary is predicted for hosts and their associated microbes. Pathways for amino acid synthesis are shared between pea aphids (Acythrosiphon pisum ) and their obligate symbiont Buchnera aphidicola (Wilson et al. , 2010). Similar interchange is predicted between the glassy winged sharpshooter (Homalodisca coagulata) and its two obligate bacterial symbionts, Sulcia muelleri and Baumannia cicadellinicola (Wu et al., 2006). Study of insect- symbiont associations can also provide insight into how animals and symbionts establish associations in the presence of immune systems meant to clear microbes and how hosts regulate symbiont populations. This list is meant to highlight how the I5K initiative can provide insight into one important aspect of insect ecology and evolution, and will hopefully inform species selection as the project proceeds. While many insects have associations with microbial partners, we focus this list on species with obligate symbiotic associations. Many of the symbiotic part ners of these insects have already been sequenced, allowing for immediate assessment of the complementarity of host and symbiont genomes. In addition to these core species that are obligate and frequently intracellular, w e also suggest a handful of species with looser symbiotic associations that can serve as a basis for beginning to explore the adaptations specific to the obligate symbiotic lifestyle. Many of these species have been nominated by other researchers for their agricultural or phylogenetic importance, highlighting the role that one species can play in shaping knowledge in many disciplines. 1 List of Suggested Focal Insects with Brief Description of Symbiotic Association s Aphi ds Cinara cedri , cedar aphid Tuberaphis styraci , gall -forming social aphid Cerataphis brasiliensis, the palm aphid Ceratovacuna nekoashi Adelges tsugae, the hemlock w ooly a delgid Daktulosphaira vitifoliae , grape phylloxera Aphids are globally distributed, economically important insects that include the first symbiotic consortia for which all members had fully sequenced genomes (IAGC, 2010; Shigenobu et al. , 2000). Currently, the genome of Acyrthosiphon pisum (pea aphid) has been sequenced, and the genomes of Myzus persicae (green peach aphid) and Aphis gossypii (cotton aphid) are being sequenced. We propose to sequence a phylogenetically nested group of aphid taxa that differ in their symbiotic consortia. These include (1) taxa that depend solely on the primary obligate intracellular gammaproteobacteria Buchnera aphidicola (A. pisum , M. persicae , A. gossypii and Ceratovacuna nekoashi ); (2) taxa dependent on a bacterial consortium (Cinara cedri, which needs both Buchnera aphidicola and Serratia symbiotica); (3) taxa that have secondarily developed novel microbial consortia (Tuberaphis styraci , a gall forming aphid in which Buchnera has been replaced by an extracellular fungal symbiont (Hongoh, Ishikawa, 2000) and Cerataphis brasiliensis , which also has a fungal rather than bacte rial symbiont) ; and (4) outgroups that include an a delgid (Adelges tsugae) that possesses a Buchnera - like symbiont and Daktulosphaira vitifoliae , grape phylloxera, an aphid- like insect that does not contain Buchnera . Many of these species can be reared in the laboratory, and many have associations with facultative symbionts as well. Mealybugs Planococcus citri , citrus mealybug The citrus mealybug, Planococcus citri , is globally distributed and feeds on phloem sap. Among other plants, it is a pest of citrus and ornamental plants. P. citri represents the scale insects, the sister taxon of aphids, phylloxerids and adelgids (Von Dohlen, Moran, 1995); thus it is an outgroup for studies on symbioses of aphids and their kin. Like aphids and other phloem feeders, P. citri relies on intracellular bacterial symbionts to provision them with essential amino acids deficient in their phloem sap diet. Unlike aphids and other phloem feeders, P. citri uniquely harbors two nested symbionts (Tremblaya and Moranella) within their bac teriocytes (von Dohlen et al. , 2001). The genomes of Tremblaya and Moranella reveal a tightly interdependent network of amino acid metabolism between the symbionts that most likely also includes the host (McCutcheon, Von Dohlen, 2011). The nested organization of these symbionts provides an opportunity to study how 2 regulation of symbionts , and metabolic interactions between the symbionts and the host, are influenced by an additional level of compartmentalization from wh at is observed in other insects. Leafhoppers Euscelidius variegatus , leafhopper The leafhopper Euscelidius variegatus does not have an association with an ancient obligate symbiosis but presence of the maternally transmitted primary symbiont BEV, which has b een sequenced, affects fecundity, longevity and developmental time. Unlike many prim ary symbionts, this symbiont persists both within tissues and freefloating in the hemolymph, suggesting that it must evade host immune responses or that the host recognizes but does not clear its beneficial partner. Facilitating future investigations, the symbiont BEV can be cultivated, allo wing for manipulation of host - symbiont associations (Degnan et al. , 2011). True Bugs Megacopta cribraria , soybean bug or kudzu bug Riptortus clavatus , broad -headed bug Megalotomus quinquespinosus , lupine bug The stinkbug Megacopta cribraria is a pest on soybean s in ma n y parts of Asia and has recently invaded North America (Jenkins et al. , 2010), where it is spreading rapidly in the Southeastern United States. The bugs are born symbiont free and then immediately feed on symbiont housing capsules left near the eggs by their mothers (Hosokawa et al. , 2007a). This allows for easy comparison of symbiont- free and symbiont- harboring insects. Work on Japanese populations suggests that the symbiont genotype dictates whether a Megacopta can or cannot use soybeans (Hosokawa et al. , 2007b). The symbiont genome has been sequenced. M. cribraria can be reared in the lab. The holy grail of studying host- microbe interactions is to be able to work wit h experimentally tractable systems in which host and symbiont can be reared separately, allowing for (1) easy comparison of symbiont and symbiont - free hosts, and (2) genetic manipulation and then reintroduction of symbionts. Unlike most insects, which pass their obligate symbionts vertically from one generation to the next, broad - headed bugs acquire their symbiotic Burkholderia bacteria from the environment in every generation (Kikuchi et al. , 2011). In at least two spec ies, Riptortus clavatus and Megalotomus quinquespinosus , the bacter ia increase developmental rate and adult size (Kikuchi et al. , 2007)(N.M. Gerardo, unpublished data) . E nvironmental acquisition provides several benefits for the study of symbiosis. First, because the bacteria must survive outside the host, they can be cultivated, facilitating genetic manipulations of the symbiont and controlled infections of the host. Second, symbiotic and aposymbiotic insects can be ea sily reared and compared. This model is an excellent parallel to the well - studied symbioses of Vibrio bacteria and squid , and of rhizobial bacteria and legume plants , 3 which also acquire their bacterial symbionts from the environment. Because of their potential as models for study of host- symbiont interactions, w e suggest sequencing of one Asian (Riptortus clavatus ) and one North American (Megalotomus quinquespinosus ) host, each of which are amenable to laboratory rearing. Cicadas, Sharpshooters and Spittlebug s ( Auchenorrhyncha) Diceroprocta semicincta , cicada Homalodisca vitripennis , g lassy-winged sharpshooter Clastoptera arizonana, spittlebug X ylem - feeding insects of the Auchenorrhyncha are sustained by symbiotic consortia centered around Sulcia mu e lleri (a Bac teroidetes) and partnered with different coresident symbionts from very different bacterial taxa. In all cases Sulcia synthesizes the majority of the essential amino acids while the bacterial coresidents synthesize the remaining minority; the amino acid biosynthetic capabilities of the coresident symbionts are perfectly complementary (McCutcheon, Moran, 2010). Based on the capabilities of the bacteria, each host is predicted to play a role in transport and synthesis. Genome sequencing of Diceroprocta semicincta, Homalodisca vitripennis and Clastoptera arizonana will facilitate confirmation of these predictions. Additionally, H. vitripennis is a major vector of the bacterial plant pathogen Xylella fastidisoa, which is the cause of variety of plant diseases, including Pierce’s disease in grapes and phony peach disease. Beetles Lasioderma se rricorne , cigarette beetle Stegobium paniceum , drug store beetle Dendroctonus ponderosae , mountain pine beetle While many of the best - studied insect- microbe symbioses involve bacteria, several insects have fungal rather than bacterial partners. The complexity of fungal eukaryotic genomes may fundamentally change selection pressure s on the genome of their insect hosts relative to selection on insects with bacterial partners . Lasioderma serricorne and Stegobium paniceum have fungal endosymbionts (Symbiotaphrina kochii and S. buchneri , respectively) that live inside their gut (Noda, Kodama, 1996). In contrast, Dendroctonus ponderosae has a suite of ectosymbionts that they use to line their broad chambers . The genome of one of these fungal symbionts ( Grosmannia clavigera) has been sequenced (DiGuistini et al., 2011). Sequencing of these insects would provide insight into three pests and would facilitate exploration of how fungal vs. bacterial symbionts, and how endosymbionts vs. ectosymbionts, impact host genome evolution. Termites Macrotermes bellicosus , African fungus -growing termite Genome sequences of two fungus- growing ants ( Atta cephalotes and Acromyrmex octospinosus ) have revealed the potential for genome complementarity between 4 insects and exosymbionts (i.e., symbionts that live outside of the host) (Suen et al. , 2011). Specifically, both ants are missing part of the arginine synthesis pathway, and it is hypothesized that this loss is compensated for by the fungi that the ants’ cultivate . Sequencing of fungus - growing termites will provide confirmation of such complementarity in a different insect lineage. Sequencing of one fungus- growing termite, Macrotermes natalensis is underway, and we suggest sequencing of other fungus - growers, including Macrotermes bellicosus . Categorization of Insect -Symbiont Systems in Terms of Key Features This list is not meant to be exhaustive but highlights some of the key features of potentially informative hosts of obligate symbioses. Some species are listed more than once. Species with sequenced symbiont genomes but no available host genome • Megacopta cribraria , soybean bug • Cerataphis brasiliensis, palm aphid ( sequencing of fungal symbiont underway) • Ceratovacuna nekoashi • Cinara cedri , cedar aphid • Planococus citri , the citrus m ealybug • Draeculacephala minerva , green sharpshooter • Homalodisca vitripennis, glassy - winged sharpshooter • Clastoptera arizonana, spittlebug • Dendroctonus ponderosae , mountain pine beetle • Diceroprocta semicincta , desert cicada • Pachypsylla venusta , hackberry petiole gall psyllid Species with easily manipulated symbiotic associations • Megacopta cribraria , soybean bug (symbiont can be removed before birth) • Riptor t us clavatus, b road- headed bug (bacteria can be cultivated) • Megalotomus quinquespinosus, broad- headed bug (bacteria can be cultivated) • Euscelidius variegatus , leafhopper (bacteria can be cultivated) • Glossiana morsitans, t setse fly (bacteria can be genetically manipulated) • Rhodnius prolixus, kissing or assassin bug (bacteria can be genetically manipulated) Species with unusual or atypical symbiotic associations • Tuberaphis styraci (aphid with fungal rather than bacterial symbiont) • Cerataphis brasiliensis (aphid with fungal rather than bacterial symbiont) • Riptorus clavatus , b road- headed bug (environmentally acquire symbiont every generation) • Megalotomus quinquespinosus , broad- headed bug (environmentally acquire symbiont every generation) 5 • Planococcus citri (one bacterial symbiont lives inside the other) Species that serve as o utgroups to those with well -studied obligate symbioses • Adelges tsugae, the hemlock wooly adelgid • Daktulosphaira vitifoliae, grape phylloxera • Planococus citri , the citrus mealybug Species currently proposed for sequencing via I5K that could inform studies of symbioses (as of Aug 1 2011) * • Anasa tristis, the squash bug • Attine ants, fungus growing ants (multiple species proposed) • Cicadulina mbila, maize leafhopper • Cimex lectularius, bed bug • Dendroctonus spp. , pine beetle s (multiple species proposed) • Euryaulax carnifex, sugarcane spittlebug • Euscelidius variegatus, leafhopper • Frankliniella occidentalis, western flower thrip • Halyomorpha halys, brown marmorated stink bug • Homolodisca vitripennis, glassy - winged sharpshooter • Lygus lineolaris, tarnished plant bug • Macrosteles quadrilineatus, aster leafhopper • Nilaparvata lugens, brown planthopper • Oncopeltus fasciatus, large milkweed bug • Pemphigus obesinymphae, popular petiole gall aphid • Planococcus citri, citrus mealybug • Rhopalosiphum padi, bird cheery- oat bug • S caphoideus titanus, American grapevine leafhopper • Schizaphis graminum, greenbug * This list is not exhaustive and will likely expand as the number of proposed species expands. Completed or c urren t i nsect genome projects with a ssociated o bligate symbioses • Acythrosiphon pisum , pea aphid • Aphis gossypi , cotton aphid • Camponutus floridanus, carpenter ant • Glossian morsitans, tsetse fly • Macrotermes natalensis, fungus - growing termite • Myzus persicae, green peach aphid • Pediculus humanus , body louse • Atta cephalotes, fungus - growing ant • Acromyrmex octospinosus , fungus - growing ant • Rhodnius prolixus, kissing or assassin bug 6 References Degnan PH, Bittleston LS, Hansen AK, Sabree ZL, Moran NA, Almeida RPP (2011) Origin and Examination of a Leafhopper Facultative Endosymbiont. Current Microbiology 62 , 15651572 DiGuistini S, Wang Y, Liao NY, Taylor G, Tanguay P, Feau N, Henrissat B, Chan S K, Hesse- Orce U, Alamouti SM, Tsui CKM, Docking RT, Levasseur A, Haridas S, Robertson G, Birol I, Holt RA, Marra MA, Hamelin RC, Hirst M, Jones SJM, Bohlmann J, Breuil C (2011) Genome and transcriptome analyses of the mountain pine beetle - fungal symbiont Grosmannia clavigera, a lodgepole pine pathogen. Proceedings of the National Academy of Sciences of the United States of America 108 , 2504- 2509 Gerardo NM, Wilson ACC (2011) The power of paired genomes. Molecular Ecology 20 , 2038- 2040 Hongoh Y, Ishikawa H (2000) Evolutionary studies on uricases of fungal endosymbionts of aphids and planthoppers. Journal of Molecular Evolution 51 , 265- 277 Hosokawa T, Kikuchi Y, Fukatsu T (2007a) How many symbionts are provided by mothers, acquired by offspring, and needed for successful vertical transmission in an obligate insect - bacterium mutualism? Molecular Ecology 16 , 5316- 5325 Hosokawa T, Kikuchi Y, Shimada M, Fukatsu T (2007b) Obligate symbiont involved in pest status of host insect. Proceedings of the Royal Society B -Biological Sciences 274 , 19791984 IAGC (2010) Genome sequence of the pea aphid Acyrthosiphon pisum . PLoS Biology 8 , e1000313 Jenkins TM, Eaton TD, Suiter DR, Eger JE, Ames LM, Buntin GD (2010) Preliminary genetic analysis of a recently - discovered invasive true bug (Hemiptera: Heteroptera: Plataspidae) and its bacterial endosymbiont in Georgia, USA. Journal of Entomological Science 45 , 62- 63 Kikuchi Y, Hosokawa T, Fukatsu T (2007) Insect - microbe mutualism without vertical transmission: a stinkbug acquires a beneficial gut symbiont from the environment every generation. Applied and Environmental Microbiology 73 , 4308- 4316 Kikuchi Y, Hosokawa T, Fukatsu T (2011) An ancient but promiscuous host - symbiont association between Burkholderia gut s ymbionts and their heteropteran hosts. Isme Journal 5 , 446- 460 McCutcheon JP, Moran NA (2010) Functional convergence in reduced genomes of bacterial symbionts spanning 200 My of evolution. Genome Biology and Evolution 2 , 708- 718 McCutcheon JP, Von Dohlen CD (2011) An interdependent metabolic patchwork in the nested three- way symbiosis of mealybugs. Current Biology in press Moran NA (2006) Symbiosis. Current Biology 16 , R866- R871 Noda H, Kodama K (1996) Phylogenetic position of yeastlike endosymbionts of Anobiid beetles. Applied and Environmental Microbiology 62 , 162- 167 7 Shigenobu S, Watanabe H, Hattori M, Sakaki Y, Ishikawa H (2000) Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS. Nature 407 , 81- 86 Suen G, Teiling C, Li L, Holt C, Abouheif E, Bornberg- Bauer E, Bouffard P, Caldera EJ, Cash E, Cavanaugh A, Denas O, Elhaik E, Fave MJ, Gadau JR, Gibson JD, Graur D, Grubbs KJ, Hagen DE, Harkins TT, Helmkampf M, Hu H, Johnson BR, Kim J, Marsh SE, Moeller JA, Munoz - Torres MC, Murphy MC, Naughton MC, Nigam S, Overson R, Rajakumar R, Reese JT, Sco tt JJ, Smith CR, Tao S, Tsutsui ND, Viljakainen L, Wissler L, Yandell MD, Zimmer F, Taylor J, Slater SC, Clifton SW, Warren WC, Elsik CG, Smith CD, Weinstock GM, Gerardo NM, Currie CR (2011) The genome sequence of the leaf- cutter ant Atta cephalotes reveals insights into its obligate symbiotic lifestyle. PLoS Genetics 7 von Dohlen CD, Kohler S, Alsop ST, McManus WR (2001) Mealybug beta - proteobacterial endosymbionts contain gamma - proteobacterial symbionts. Nature 412 , 433- 436 Von Dohlen CD, Moran NA (1995) Molecular phylogeny of the Homptera: a paraphyletic taxon. Journal of Molecular Evolution 41 , 211- 223 Wilson ACC, Ashton PD, Calevro F, Charles H, Colella S, Febvay G, Jander G, Kushlan PF, Macdonald SJ, Schwartz JF, Thomas GH, Douglas AE (2010) Genomic insight into the amino acid relations of the pea aphid Acyrthosiphon pisum with its symbiotic bacterium Buchnera aphidicola . Insect Molecular Biology 19 , 249- 258 Wu D, Daugherty SC, Van Aken SE, Pai GH, Watkins KL, Khouri H, Tallon LJ, Zaborsky JM, Dunbar HE, Tran PL, Moran NA, Eisen JA (2006) Metabolic complementarity and genomics of the dual bacterial symbiosis of sharpshooters. PLoS Biology 4 , 1079- 1092 8