UNIT 1: INTRODUCTION TO BIOLOGY
advertisement

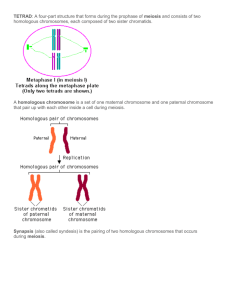
SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. Chapter 8: Reproduction, Meiosis & Genetic variation - Part II Reproduction and the role of germ cells Reproduction is one of the hallmark characteristics of all forms of life on planet Earth all living organisms reproduce; that means during reproduction they hand over (= inherit) their individual genetic make-up and information over to a next generation which assures the continuation of a species reproduction (in biological terms) is a hallmark feature of living beings and means the formation of a new daughter organism from a pre-existing parental one - for most organisms the new individual starts with a fertilized egg in a process called fertilization and a series of subsequent cell divisions two major forms of reproduction are established in the living world 11.. S Seexxuuaall rreepprroodduuccttiioonn - it is the predominant form of reproduction of m muullttii--cceelllluullaarr eeuukkaarryyootteess and aanniim maallss - on the macroscopic anatomical level, sexual reproduction means the interaction of two individuals of one species with a different gender (male or female) with the biological purpose to create new offspring - on the molecular and cellular level it means the merging of genetic material in form of chromosomal DNA after fusion of one female-derived egg cell and one male-produced sperm - after successful fusion of one haploid sperm with one haploid oocyte (= egg cell) in a biological process called fertilization, a diploid, so-called zygote forms which starts the life cycle of a new individual (see Figure of life cycle of humans below) - the haploid or single chromosomal set of genetic material from each parent is fused to create a new individual with a so-called diploid gene-set Sex and sexual reproduction from a biological perspective As humans we may have asked ourselves the same simple question at some point of our life. Why are there two separate genders established amongst humans and not more … or just one? How come that not every creature on this planet does a living as a hermaphrodite, comprising two genders in one body? Why is there two sexes established in most higher evolved members of the living world and how come that these two sexes of a given species somehow desire to get involved with the other gender of the same species to perform acts for the purpose of sexual reproduction? Well, we easily agree that sex and sexual reproduction is responsible for most what is flamboyant, colorful and beautiful in nature, i.e. the colorful showy flowers of plants, the melodies of bird songs, poetry and arts, it can’t be the sole purpose of it. Biologists agree today, that all these practices, performances and show-offs associated with the phenomenon of sexual reproduction are just the means to an end which is sexual reproduction as an effective means to create an increased survival chance of a species in a changing world due to increased genetic variability. SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. In order to survive as a species in an ever changing world, the species has to change in morphology and behavior over time to be able to meet the new environmental challenges. Since molecular biology tells us today that DNA, genes and genetic programs dictate the outer appearance and morphology of living organisms, species can only change if they change their genetic make-up. No genetic variation…no evolutionary change of life on this planet! That is exactly where sexual reproduction and the molecular process called meiosis kicks in. As you will see in the following sections below, sexual reproduction and the connected biological process meiosis assure the genetic material becomes shuffled and shuffled again during each generational cycle to purposely increase the genetic variability of that very species. 22.. A Asseexxuuaall rreepprroodduuccttiioonn - this form of reproduction is observed mostly in bacteria, amoeba and plants in the plant kingdom asexual reproduction leads to the quick formation of certain plant structures, such as “runners”, in usually stable environments - bacteria reproduce asexually in process called ‘binary fission’ , which is the “pinching-off” of one parent cell into two new bacterial cells (see Image below) - the production of new offspring by a single parent organism - DNA of the offspring is inherited from one parent or cell and is identical with the parental DNA Binary fission of bacteria Rod-Shaped Bacterium, hemorrhagic E. coli, strain 0157:H7 (division) (SEM x22,810) In the life cycles of sexually reproducing organisms we observe an alternation between a haploid stage of the organism, which is characterized by cells with a single chromosome set, and a diploid stage (see Graphic below), which cells show a double chromosome set SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. Alternation of haploid (1n) and diploid (2n) stages in the human life cycle Gonads Ovaries Testis Zygote ( 2n ) Gametes (1n ) Oocyte (egg cell) Sperm 23 chromosomes ( = 1n ) 23 chromosomes ( = 1n ) SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. 33.. P Paarrtthheennooggeenneessiiss - This rare form of reproduction is performed by cceerrttaaiinn aam mpphhiibbiiaannss aanndd rreeppttiilleess - the females of these species are able (dependent on environmental factors) to switch between sexual and asexual reproduction - a very prominent example for this kind of reproduction is the whiptail lizard which lives in the Southwestern part of the U.S. TThhee rroollee ooff ggeerrm m cceellllss iinn rreepprroodduuccttiioonn While mitotic cell division (see Chapter 8) can be performed by literally all somatic cells in a multi-cellular organism, formation of reproductive cells, or so-called sex cells (or gametes), is only performed by a unique class of germ cells in sexually reproducing organisms, the so-called gamete mother cells or primordial germ cells The germ cells are located within the sex cell-producing organs or gonads, i.e. testes (male gender) or ovaries (female gender), of sexually reproducing organisms Within the gonads, the primordial germ cells perform a unique form of cell division, called meiosis, which is in some parts similar to mitosis but shows major and crucial differences to mitotic cell division at some stages One important difference to mitosis is the interesting fact that meiosis does not lead to daughter cells with the same amount of chromosomes as the parental cell; the daughter cells, i.e. gametes, resulting from meiosis have only half the chromosome number as the parent cell “ Meiosis leads to a 50% reduction of the number of chromosomes in the resulting daughter cells …” As you remember form the previous chapter, the total cellular DNA of a species (= genome) is split into a certain number of chromosomes the number of chromosomes is unique for each species; different species have different number of chromosomes each cell of the human body, or so-called somatic cell, has 46 chromosomes in its nucleus; the only exception are the so-called gametes (= sperm and egg cells) which have only 23 chromosomes it is these gametes (or sex cells) in biological organisms which are produced by earlier mentioned so-called germ cells in humans and in higher organisms the chromosomes in each somatic cell occur in pairs or sets of two; each cell which has a double set of chromosomes is called diploid (= 2n) each set of chromosomes was received from the biological parents during Fertilization SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. simple organisms or the gametes (eggs or sperm) of higher organisms have only one set of chromosomes; they are haploid (= 1n) since most species, including humans, have two sets of chromosomes; they are also called diploid (= 2n) organisms in diploid organisms one set of chromosomes comes from the egg cell of the mother, the other set of chromosomes comes from the sperm of the father the pairs of identically shaped chromosomes within the somatic cells of diploid organisms are also called the homologous chromosomes the characteristics of homologous chromosomes are: 1. each pair of homologous chromosomes contains its genes at the same place or so-called locus on the DNA double helix 2. a gene on the identical locus on a homologous chromosome codes for the same inherited characteristic, e.g. eye color, blood group 3. but the genes on the corresponding loci on the chromosomes are slighly different, i.e. have a variation its DNA nucleotide sequence; biologists say the gene exists in two different alleles of the 23 pairs of homologous chromosomes in humans, 22 pairs are chromosomes called autosomes autosomes are found in cells of males and females - the other pair of chromosomes is called the sex or gender chromosomes the genes found on it determine a persons or species gender all human females have a pair of X-chromosomes (= XX genotype) while males in contrary have one X and one significantly smaller sized and differently shaped Y-chromosome Gender chromosomes of other biological organisms & Gender determination While the XX genotype in humans leads to the expression of female body features and the XY combination determines maleness in humans, gender determination in other organisms can be completely different. Indeed, biologists found a series of interesting variations to this theme in nature. For example, lemmings have three sex chromosomes, called W, X and Y. In these truly interesting mammals, the XY genotype leads to male animals, while any of the alternative chromosomal combinations XX, WX or WY, leads to femaleness. The also possible YY genotype is not compatible with life and these sons die. The Australian billed, egg-laying mammal Platypus has been shown to have 5 pairs of sex chromosomes which determine the gender in this species; the XXXXXXXXXX genotype is found in the female gender of this species, while males have five XY pairs (= XYXYXYXYXY) in their cells! SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. In birds, the story is upside down. The XY or only X genotype leads to female birds, while maleness is genetically dictated by the XX genotype. Most peculiarly, some animals don’t have sex chromosomes at all. TThhee ddoouubbllee cchhrroom moossoom mee sseett ooff aa hhuum maann ssoom maattiicc cceellll 1 – 22: autosomes gender or sex chromosomes 2 sets of chromosomes is a hallmark of cell nuclei of higher animals and humans the complete set of chromosomes of an organism is called the diploid number the diploid number is species-dependent; it varies from species to species - e.g. the diploid number for humans is 46 - the ant Myrmecia pilosula has a diploid number of 2; the females have only a single pair of chromosomes; the males of this group of ants develop from unfertilized (haploid) eggs and, hence, each of their cells, has only one single chromosome!! SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. - the extremely polyploid fern Ophioglossum reticulatum has a diploid number of 630 (!!), which means that there are 1260 chromosomes per cell summarized, all cells of the human body are diploid with the exception are the so-called sex cells or gametes the germ cells of ffeem maallee organisms are called oocytes (or egg cells) the m maallee gametes are called ssppeerrm m cells gametes have only have one single set of chromosomes in their nucleus; we say these cells are haploid the human life cycle starts with the fusion of two haploid cells (= the female oocyte with the male sperm cell) in a process called fertilization (see Figure above) after fertilization the diploid fertilized egg or also called zygote starts to divide by continuous mitosis to form the diploid embryo & fetus early during embryogenesis, haploid gametes are formed by specialized cells, the socalled germ line cells, by a unique form of cell division called meiosis, which individual events and phases we will look up in the following sections in more detail MEIOSIS … creates genetic variety Meiosis is a special type of cell division in which DNA replication is followed by to rounds of chromosome segregation; homologous chromosomes segregate during the first and sister chromatids split in the second division; as a consequence, meiosis creates: 1. haploid germ cells (eggs and sperm) from a diploid parent cell for sexual reproduction 2. genetic variety due to tetrad formation and crossing over O Onnee single diploid parent cell is divided to produce ffoouurr haploid daughter cells (= gametes) these haploid germ cells are created in an very early stage of the female human life cycle they remain slumbering in the G2/M-phase of meiosis I for many years (= “meiotic arrest”) they grow out or also called differentiate into the functional (= mature) oocytes after onset of puberty; this differentiation process is triggered by the release of certain hormones e.g. GTRH, FSH this final maturation of the germ cells or differentiation occurs in the so-called reproductive organs or gonads of m maalleess (= testes) and ffeem maalleess (= ovaries) Meiosis involves a reduction of the genetic material from a double (= diploid, 2n) chromosomal set to a single (= haploid, 1n) set SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. Meiosis comprises two successive nuclear divisions (= Meiosis I and Meiosis II) with only one round of DNA replication in Interphase I Overview: Stages of meiosis P Prriim moorrddiiaall ggeerrm m cceellll oorr ggaam meettee m mootthheerr cceellll Gametes (egg cells or sperm) diploid haploid The two major steps of meiosis in more detail: M Meeiioossiiss II (= homologous chromosomes separate) IInntteerrpphhaassee II 1. the chromosomal DNA of the cell becomes duplicated by DNA replication 2. the resulting duplicated chromatin fibers remain attached with each other with the help of “cohesive proteins” (see cohesins in the Figure below) 3. the chromatin fibers, consisting of DNA and histone proteins, are not visible in the light microscope at this point 4. the centrosomes (including the centrioles) duplicate at this phase and form the microtubule organization center (= MTOC) of the meiotic cell P Prroopphhaassee II 1. this phase occupies approximately 90% of the time required for cell division 2. the cell initiates the heavy coiling and folding of the chromosomal material in a process called DNA condensation; as a consequence the chromosomal DNA becomes visible when observed with a light microscope - each duplicated chromatin fiber turns becomes visible in form of sister chromatid arms (see Figure below) SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. DNA condensation & Protein modifications In the past years molecular biologists figured out that DNA condensation is accompanied by the heavy modification of the histone components of the chromatin fibers. According to several studies, DNA condensation involves the molecular modification of the chromatin protein histone H3 by phosphorylation of its serine 10 residue in the N-terminal end (= serine phosphorylation). Other studies point the role of methylation of critical DNA bases (= DNA methylation) by enzymes called Methyltransferases to support compacting the DNA. 3. the synaptonemal complex (SC) forms (see Figure below) between homologous chromosomes; as a consequence the homologous chromosomes pair with each other in a process called synapsis - many proteins have been identified which are responsible for the build-up are (at least temporarily) part of the synaptonemal complex - proteins such as Scp1 (SynI/Sycpi), Scp2, Scp3 (Cor1) and Fkbp6 The synaptonemal complex & Fertility The importance of these SC proteins for proper meiosis has been shown in so-called “knockout mice” models; for example: male mice in which the gene for Scp3 has been knocked out by molecular biological techniques, or so-called Scp3-/- mice, show infertility and aneuploidy; Fkbp6-/- mice show male infertility and the testicular spermatogenesis is blocked in these animal. A better understanding of this part of meiosis may lead to future cures of infertility; according to one source, about 15% of couples worldwide remain childless due to infertility Formation of the synaptonemal complex to assure tetrad formation during meiosis Sister chromatid Tetrad formation Synaptonemal Complex (SC) Synapsis of homologous chromosome pair Synaptonemal proteins Scp1 (SynI/Sycpi) Scp2 Scp3 (Cor1) Fkbp6 Cohesin SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCE Molecular & Cellular Biology; Instructor: Elmar Schmid, Ph.D. 4. as a result of synapsis the so-called tetrads form within the cell - tetrad formation (tetra = four) means the side-by-side alignment of four sister chromatids during this phase 5. tetrad formation during Prophase facilitates the exchange of DNA segments of the neighboring sister chromatids of homologous chromosomes in a process called recombination by crossing over Crossing over & Sex chromosomes “Almost all chromosomes are equal if it comes to crossing over, except the sex or gender chromosomes …” While all human autosomes are equally likely involved in crossing over events during prophase I of meiosis, the sex or gender chromosomes are the great exception. Molecular biologists unraveled that 95% of the DNA of the human Y chromosome, which comprises 60 million base pairs and contains the genes responsible for the expression of the male phenotype, is non-recombining. Due to the fact that the genetically degenerate Y chromosome (it has lost most of the active genes that were present in their ancestor chromosomal DNA) has a very short-armed, so-called non-recombining region (= NRY region) it is not involved in recombination event with the X- chromosome by crossing over during meiosis. The genes located in the NRY region of the human Y chromosome and its variations in males with different ethnic or geographical background are therefore very attractive molecular markers and are of high scientific interest for population genetics and to unravel human history. Y-chromosomal DNA and its interspecies variation are also promising tools in modern forensic science. Crossing over of homologous chromosomes during meiosis SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. during crossing-over sister chromatids strands break and are reattached to the neighboring homologous chromosome the molecular biological events of the this amazing and important event of meiosis are still not completely understood, but scientists unraveled the involvement of several key events and molecular key players in this event (see Graphic below) According to the double-strand break model of meiotic recombination (see Graphic below), following sequence of events happen during crossing over to form chromosomes with re-combined sister chromatic DNA or so-called recombinant chromosomes 1. After synapsis and chiasma formation, one double strand break happens at one of the sister chromatid strands at the cross point; some nucleotides are removed on both strands of the cut DNA in 5’ – 3’ direction by a DNAase; DNA becomes resected in that region 2. one strand of the broken DNA (= dark blue line in Graphic below) is transferred over to the complementary region of the aligned homologous chromosome (red lines in Graphic below) which has been opened to form an “invasion loop” at the point of invasion of the strand 3. After Watson-Crick base pairing with the complementary DNA strand, new nucleotides are added to the free 3’-end of the cut strand end with the help of a DNA polymerase enzyme; one strand of the non-cut homologous chromosome serves as template during this DNA synthesis reaction; this step is also called DNA repair synthesis 4. Nucleotides are filled into the second strand of the cut chromosomal DNA (upper bright blue line in Graphic below) by DNA repair synthesis to form a double-stranded, recombinant DNA at this stage; the DNA fragments are chemically glued together (= ligated) at the previous break points with the help of an enzyme called ligase and two complex DNA tangle structures form at the former double strand break region, these peculiar DNA structures are named after its discoverer Holliday structure (see the two X points in step 4 of the Graphic below) 5. In a final step DNA is cleaved and re-ligated again at the Holliday structures to form alternative recombinant homologous chromosomes The exact molecular mechanisms and involved proteins and/or enzymes are vastly enigmatic and is still an area of intensive scientific research crossing over leads to a rearrangement (= shuffling) of the genetic material and increases the genetic variability of the genetic material in sexually reproducing organisms (see sections below!) 11 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Molecular Biology of Crossing Over in Meiosis I Double-strand break model of meiotic recombination Homologous Chromosomes (Tetrad formation) Double-strand break & 5’ end resection Strand invasion DNA synthesis (with D as template) Repair synthesis & Ligation 12 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 6. another important event of prophase I is the disappearance of the nucleoli within the cell nucleus 7. the nuclear envelope is proteolytically digested (= with the help of a unique class of enzymes called proteases); as a consequence the cell nucleus breaks into fragments and gradually disappears within the cell 8. the (previously duplicated) two centrosomes (= the structures where the later microtubules begin to grow out by polymerization) move away from the center core and slowly migrate into opposite direction towards the two cell poles 9. Chromosomes move away, fill out the complete interior cell space and the spindle apparatus forms 10. Tetrads are moved to the cell center guided by microtubules M Meettaapphhaassee II aanndd A Annaapphhaassee II 1. Chromosomal tetrads are aligned on the metaphase or equatorial plate - Tetrad alignment happens randomly at this step (Random alignment!) - That means the maternal (red color) and paternal chromosomes (blue color) of the tetrads will randomly face different cell poles (see Graphic below) - Depending on the number of homologous chromosome pairs in the meiotic cell (or the so-called haploid number n), 2n possible chromosomal alignments (= Ncomb.)arise at this step - For example,: in the case of a human meiotic cell with a haploid number n of 23, more than 8 million (!!) different chromosomal alignments are possible during this step! 13 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Random alignment of homologous chromosomes in Meiosis I General: Haploid number n=3 Ncomb = 2n DNA-replication For this example: Prophase I 3 Ncomb = 2 = 8 or or or or or or or or Metaphase I Alignments 2. Two large protein complexes form around the centromere regions of each chromosome and build the so-called sister kinetochores (see Graphic below) - interestingly, in meiosis, the sister kinetochores form on only one side of the homologous chromosome pair and show so-called “co-orientation” - recall: in mitosis the kinetochores form on opposite sides on the chromosomal centromere region 14 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Sister kinetochores of meiotic metaphase chromosomes Homologous Chromosome (tetrad) Microtubul Sister kinetochores (“co-orientation”) 3. In a next step, hollow protein fibers called microtubules, which “grew out” from the cell pole-positioned centrosomes by polymerization, become attached to one or two of the kinetochores on each of the cellular (metaphase) chromosomes - the primary function of the kinetochore complex known today is to guide in and capture the microtubule coming from one of the MTOCs located in the cell poles - proper timing and status of microtubular attachment is carefully monitored and surveilled by the cellular, so-called “spindle checkpoint system”; the latter involves an arsenal of recently discovered check point proteins, such as the MAD (MAD1, MAD2, MAD3) and BUB (BUB1, BUB3) proteins - the proteins of the meiotic check point system detect the presence of even a single unattached kinetochore and stops (“arrests”) the further progress of meiosis until the unattached kinetochore succeeded in capturing microtubules from the distant spindle pole Molecular biologists unraveled that the sister kinetochores do not capture the microtubules simultaneously, and that some kinetochores have not finished proper attachment to microtubules by the time the cell moves in the next phase of meiosis (= anaphase). As a (dangerous) result of “unattached kinetochores” during meiosis, mis- or unequal segregation of homologous chromosomes, gametes with wrong numbers of chromosomes are the outcome with usually disastrous consequences for sexual reproduction and conception (see section: “Non-disjunction during meiosis” further below. 15 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Yeast cells which have a defective meiosis check point system due to a mutated MAD2 protein show mis-segregated chromosomes after completion of meiosis. According to one study, 20% of all human conceptions have major chromosomal abnormalities caused by mis-segregation of chromosomes during meiosis, which proves the enormous importance of this meiotic step! 4. During Anaphase I, a protein complex called anaphase-promoting complex (=APC) becomes activated by cell cycle regulators, which leads to the subsequent degradation of the synaptonemal complex by a protease - even though the exact mechanism is not known yet, but scientists could show the involvement of a series of important key player proteins and enzymes in this important event, such as the enzyme Polo-like kinase CDC5, CDC20 and Pds1 The molecular biology behind homologous chromosome segregation during meiosis I - according to recent studies conducted in the budding yeast, major molecular player seems to be Pds1, a cell protein which inhibits the protease enzyme Esp1; as a consequence, Pds1 cannot cleave the cohesin subunits - cell cycle-activated CDC20 prepares the Esp1 activity blocking Pds1enzyme for proteolytic (?) degradation; Pds1 becomes active - simultaneously, CDC5 kinase phosphorylates important cohesin sub-units, such as Scc1/Med1, to support effective cohesin degradation by the activated Esp1 protease 5. The kinetochore-attached microtubules (or kinetochore microtubules) begin to shorten due to continuous depolymerization; This step is very susceptible to the intervention of so-called spindle blockers, such as the plant-derived molecules taxol, vinblastine, colchicine and benomyl (see Chapter 8, part II); spindle blockers are compounds that bind to microtubular proteins and are able to interfere with either microtubule polymerization or depolymerization; as a consequence they prevent the normal dynamics of the microtubular spindle apparatus in cells 6. Due to the firm attachment of the microtubules with the sister kinetochores, the tetrads split and the homologous chromosome pairs are pulled toward the opposite cell poles; 16 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. TTeelloopphhaassee II & &C Cyyttookkiinneessiiss 1. The separated homologous chromosomes arrive at the opposite poles of the cell and each cell pole has the haploid chromosomal set consisting of two sister chromatids 2. Chromosomes uncoil and the nuclear envelope re-appears 3. At the same time, the cell stretches and elongates due to the continued polymerization of microtubule pairs (= polar microtubules) that have not attached with the chromosomal kinetochore (see Graphic below) Different roles of kinetochore and polar microtubules during anaphase I of meiosis Kinetochore microtubules Polar microtubules = = chromosomal separation cell elongation & stretching 17 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 4. A cleavage furrow forms in the equatorial line of the elongated cell with the help of a contractile actin ring, located underneath the cell membrane; continued contraction of this actin ring due to depolymerization executes the final split of the meiotic cell into two new daughter cells and the first cell division is complete - this process is susceptible to agents which interfere with actin polymerization/depolymerization, such as cytochalasin D 5. Both newly formed cells usually enter a short period of interphase before they prepare for a second round of meiotic division but this time without chromosome duplication - in human females, where meiosis I begins already during fetal development and starts 6 weeks before birth, the resulting primary oocyte at the end of this stage goes into a very long interphase period; the also referred to meiotic arrest of the two immature oocytes last until puberty (see Graphic further below) - at the onset of puberty in the female the slumbering oocyte develops into a matured primary oocyte under the influence of the hormone FSH; it then continues with meiosis II No DNA replication occurs before starting meiosis II! M Meeiioossiiss IIII (= sister chromatids separate) The cellular processes of meiosis II are essentially the same as in mitosis (for molecular details I therefore refer to part I (Mitosis) of this chapter) P Prroopphhaassee IIII 1. Chromosomes condense again 2. The nuclear envelope breaks apart again and the cell nucleus disappears within the cell 3. The spindle apparatus forms again and the 2 sister chromatids move to the middle of the cell M Meettaapphhaassee IIII aanndd A Annaapphhaassee IIII 1. The visible metaphase chromosomes are aligned in the cellular center area (= metaphase plate), with the kinetochores of each sister chromatid pointing towards opposite poles (“opposite orientation”) (see Graphic below) 2. Microtubules, arriving from the cell pole located centrosomes (or MTOCs), make contact with the opposite kinetochores of the sister chromatids; the microtubular spindle apparatus forms within the cell 18 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Opposite orientation of kinetochore during metaphase II Loss of sister chromatid arm cohesion Microtubule Cohesin Kinetochores (“opposite orientation”) 3. After cell cycle-dependent activation of the anaphase-promoting complex (APC) and other protein factors, such as cdc5/polo kinase, activated proteases, most importantly separase, begin the degrade the cohesin proteins which hold the two sister chromatids together (see Graphic below) - important roles play the anaphase inhibitor, which (in non-meiotic cells) is bound to the separase-inhibiting protein securin - after modification of the separase-inhibiting securin-anaphase inhibitor complex (SAIC) by activated APC through a protein modifying process, called polyubiquitination (Ubi), the SAIC is degraded by proteolysis within a “cellular garbage can” called proteasome 19 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. - - activated separase enzyme cleaves the cohesin proteins (which hold the two sister chromatids together) by proteolysis; the sister chromatids are now only attached to the microtubules via the kinetochores another major role in sister chromatid separation seems to play the protein kinase enzyme Polo/Cdk5, which phosphorylates the cohesin protein component Scc1 on critical serine residues 4. the freed sister chromatids are pulled in the opposite direction towards the cell poles under the force of the shortening kinetochore microtubules - microtubule shortening happens due to depolymerization of the protein strands into its tubulin monemers 5. The (now) individual daughter chromosomes move towards opposite poles along the spindle apparatus Molecular Biology of Sister Chromatid Separation during Anaphase II of Meiosis Cohesin (MPC) - Survivin - CENP-A Esp1 protease (yeast) Cell Cycle Polo/Cdk5p + Scc1 (Hs) Sec1p (Sc) Serine Phosphorylation of Scc1 Kinetochore Separase Microtubule Microtubule Sister chromatids Proteolytic Cleavage + Binding + Inhibition Securin Anaphase Inhibitor Proteasome Ubi Ubi Ubi Ubi Ubi Ubi Ubi Ubi Ubi PolyUbiquitination APC Graphics©E.Schmid/2002 20 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. TTeelloopphhaassee IIII & & ccyyttookkiinneessiiss 1. after arrival of the separated sister chromatids at the cell poles, the cell nucleus forms at each pole of the cell; the cell also activates the cellular processes necessary for successful cytokinesis and cell division, most importantly formation of the cleavage furrow with the help of a constricting sub-membranuous actin ring 2. at the end of meiosis II, 4 daughter cells are formed each with a haploid number of chromosomes (= the former sister chromatid) 21 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Meiosis: Overview of the two meiotic phases and the major cellular events Activation by FSH Diploid gamete mother cell DNA-replication Prophase I Metaphase I Anaphase I Tetrad formation & Crossing over Random alignment of homologous chromosomes Homologous chromosomes segregate Prophase II Anaphase II Sister chromatids separate Haploid gametes (sperm or oocytes) 22 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. S Suum mm maarryy:: H Haallllm maarrkk ffeeaattuurreess ooff m meeiioossiiss 1. two consecutive cell divisions but only one duplication of the chromosomes 2. four haploid daughter (= germ) cells result 3. increase of genetic variability H Hoow w ddooeess m meeiioossiiss iinnccrreeaassee tthhee ggeenneettiicc vvaarriiaabbiilliittyy iinn ooffffsspprriinngg?? the gene shuffling and recombination events occurring during meiosis creates increased genetic variability, which is the ultimate raw material for the evolutionary process of natural selection new gene combinations eventually confer novel structural, physiological or behavioral characteristics to an individual with certain advantages in a changed environment or in environments with new challenges the increase in genetic variability during meiosis is achieved by three events 1. Independent orientation of the homologous chromosomes during Metaphase I the orientation of homologous chromosomes to form a tetrad is a pure by-chance incident all homologous chromosomal pairs orient independently at Metaphase I for any species the total number Nt of possible combinations of chromosomes that are distributed into gametes is: Nt = 2nnn nn = haploid chromosome number for hhuum maannss with a haploid chromosome number of 2233, the number of possible chromosomal combinations in a gamete is: 2222333 or 8 million (!!) that means every gamete (oocyte or sperm) of humans contains one of about 8 million possible combinations of maternal and fraternal chromosomes!! 23 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 2. Random fertilization - - fertilization = the biological process during which a haploid (1n) gamete (= ooooccyyttee) from one female individual unites (= fuses) with a haploid (1n) gamete (= ssppeerrm m) from another male individual to form a diploid (2n) zygote this process is random = it is not predictable which gamete will finally fuse with which gamete but the number of possibilities of chromosomal combinations in the new diploid can be calculated it is the product of the number of chromosomal combinations of each individual e.g. for hhuum maannss, a m maann and a w woom maann can produce a diploid zygote with any of: 88 m miilllliioonnss X 88 m miilllliioonnss = 64 trillion (!!) different chromosomal combinations !! 3. crossing over Electron microscopic picture of a crossing over event crossing over is the increase of genetic variability on the gene level the exchange of corresponding DNA segments between two homologous chromosomes during Prophase I of meiosis the end of the corresponding sister chromatids of the tetrads occasionally cross over during synapsis with crossing over, new kinds of gametes occur after meiosis which harbor a new set of haploid chromosomes, the so-called recombinant chromosomes one single crossing over event can affect many genes and multiple crossing over events can in a cell considering that there are more than 1000 genes located in one side arm of a sister chromatid, any crossing over event affects and rearranges many genes, which are ‘bulk’-translocated 24 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Nobody is perfect! Meiosis neither Non-disjunctions during meiosis, chromosomal aberrations & abnormalities since meiosis consists of many intricate steps, involving many enzymes and protein components, it is a highly vulnerable cellular process which is prone to errors and can be affected by many factors mistakes or errors which affect the separation of chromosomes during meiosis I or II lead to alterations of chromosome numbers or so-called chromosomal aberrations possible accidents or malfunctions during meiosis which account for alterations of chromosome numbers are: 1. Non-disjunction in meiosis I members of the homologous chromosome pairs fail to separate during Metaphase I of meiosis (see Figure below) as a consequence n+1 or n-1 gametes form at the end of meiosis Aberrant chromosome numbers in gametes due to a non-disjunction event in meiosis I Disturbing Factor Interpha s e Propha s e I M e ta pha s e I Prop ha s e II Graphic©E.Schmid/2001 M e ta pha s e II n+1 or n-1 Gametes 25 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 2. Non-disjunction in meiosis II sister chromatids fail to separate equally during Metaphase II of meiosis (see Figure below) as a consequence, either gametes with a normal set of chromosomes are the result (n) or gametes with n+1 or n-1 numbers of chromosomes appear Normal or aberrant chromosome numbers in gametes after a non-disjunction event in meiosis II Disturbing Factor Interpha s e Propha s e I M e ta pha s e I Prop ha s e II Graphic©E.Schmid/SWC2001 M e ta pha s e II n, n+1 or n-1 Gametes fertilization involving gametes with wrong (= aberrant) chromosome numbers or chromosome patterns results in offspring with chromosomal abnormalities, = additional/extra or missing chromosomes in the resulting zygote and embryo errors of, or interference with meiosis can also lead to alterations in chromosome structures (e.g. deletions or translocations) in the gametes; e.g. mistakes happening during Prophase I of meiosis due to unequal crossing over events can (if they remain undetected by the cell’s surveillance and repair system) lead to alterations in chromosome structures in the gametes Finally, more subtle changes in chromosome structures and shapes can also be caused by a series of other factors, such as strong irradiation (e.g. X-rays, radioactivity) and environmental factors (e.g. toxins, viruses) 26 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Known events which lead to abnormal chromosomal structures are so-called: 1. Deletions a chromosomal piece breaks off and gets lost 2. Duplications a section of a chromosome duplicates and reinserts (= dup) at the same chromosome often referred to as “partial trisomy” 3. Insertions chromosomal pieces/fragments insert into other chromosomal sites 4. Inversions chromosomal piece breaks off a chromosomal site and re-inserts in opposite direction at same site 27 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 5. Translocations: - a chromosomal fragment excises and moves over to a non-homologous chromosome where it inserts - translocation in somatic cells can contribute to the development of cancer - e.g. in the cancerous white blood cells of patients suffering from chronic myelogenic leukemia (CML); in CML a translocation occurred which carried a part of chromosome 22 over to chromosome 9 and vice versa; as a result a so-called fusion protein, called Bcr-Abl is formed which leads to a dysregulation of cellular processes - a translocation between chromosomes 8 and 14 is found in most cases of a form of blood cancer called Burkitt's lymphoma Translocation of a segment of the human chromosome 5 onto chromosome 18 Chr. #: 5 18 5 5,18 28 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 5. Chromosome rings: chromosomal ends of the short p and long q arm break off and get lost the remaining chromosome ends fuse and form typical chromosomal ring structures often observed in cells after exposure to strong Xray irradiation or radioactivity which is often accompanied with DNA double strand breaks (DSBs) fertilization involving gametes with wrong (= aberrant) chromosome numbers, aberrant chromosome patterns or structures results in offspring with chromosomal abnormalities which in many cases are the cause for health disorders and clinical symptoms the most common chromosomal aberrations observed in sexually reproducing organisms are monosomies (= cells with a missing chromosome) and trisomies (= cells with an extra chromosome) severe chromosomal aberrations or defects in an individual or embyo can be early detected and analyzed by preparing a so-called karyotype 29 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Karyotype analysis Genetic disorders caused by chromosomal aberrations or defects can be detected with by performing a karyotype analysis Many genetic disorders can be even detected by karyotyping long before birth by fetal testing with the help of different clinical procedures and techniques The most commonly applied fetal testing techniques in hospitals and cytogenetic clinics are: 1. Amniocentesis - this procedure is performed on pregnant women between the 14th and 16th week of pregnancy - a sample (2mL) of the amniotic fluid is consent-collected in a hospital and biochemical tests as well as karyotyping are performed with the sample -- the complication rate leading to an unwanted abortion of the embryo is about 1-2% 2. Chorionic villus sampling (CVS) - performed between the 8th and 10th week of pregnancy - a small amount of embryonic tissue, the so-called chorionic villi, are taken from the placenta for testing - this procedure is faster due to the rapid growth of the embryonic cells of the villi Both medical procedures are usually reserved for situations in which the possibility of genetic disorders is significantly increased, such as in: 1. a 35 year old or older women with their first pregnancy 2. couples with a proven family history for a certain hereditary disorder a karyotype analysis is usually performed with isolated white blood cells (Adults) or sampled embryonic cells from the amniotic fluid or chorionic villi of the placenta; karyotyping is a lab-performed, multi-step procedure which involves following steps (see Figure below): 1. the sampled and washed cells are incubated with a potent mitogen, e.g. PHA or concanavalin A (ConA) to induce mitosis in the cultivated cells 2. the cells are blocked in Metaphase of mitosis with the help of added molecules = spindle blockers, such as colchicines, which interfere with proper microtubule formation 3. the cells are fixed, the condensed Metaphase chromosomes are stained with a certain dye (= Giemsa) and observed under a light microscope 30 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. The different steps of the karyotyping procedure Amniocentesis or Venipuncture (peripheral blood) KCl (hypo-osmotic shock) Embryonic cells or Lymphocytes Colchicine (= Mitosis blocker) PH A ConA (= Mitosis stimulus) Centrifuge Cell culture (ca. 72h) (ca. 2h) (ca. 20 min) Medium Centrifuge Pipette transfer Heating & Drying Microscope glass slide Centrifuge Fixation Giemsa dye Methyl alcohol/ Acetic Acid Mix (3:1) Stained & fixed mitotic cells Chromosome staining Graphics©E.Schmid/200 Microscopy (250x) 31 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 4. a photo is taken of the observed chromosome smear (see Figure below) Human metaphase chromosomes before establishing a karyogram 5. the observed metaphase chromosomes of the documented chromosome smear are arranged according to size, banding pattern and centromere location, in a typical karyogram (see Image below) and the karyogram carefully analyzed for chromosome abnormalities, such as trisomies, deletions or translocations a karyotype analysis involves comparing chromosomes for their: 1. 2. 3. 4. number (e.g. Trisomies) length (e.g. deletions, translocations) placement of centromeres location and sizes of chromosomal G-bands (deletions, insertions, inversions) the Giemsa dye stains regions of chromosomes that are rich in the base pairs Adenine (A) and Thymine (T) where it produces the typical dark bands, the so-called G-bands the biomedicinal process leading to a karyotype is called karyotyping during karyotyping, the homologous chromosomes are arranged in a so-called karyogram and compared “Every year over 400,000 karyotype analyses are preformed in the U.S. and Canada in the genetic labs of hospitals or clinics.” The graphic below shows a typical karyogram of a human female after successful karyotyping - the homologous chromosomes are identified, cut out and arranged according to size and chromosome number - chromosome #1 is the largest human chromosome, while the small chromosome #22 is shown second last in this typical karyogram below 32 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. Karyogram of a human female 1 2 8 9 16 17 3 10 18 Autosomes 11 19 4 5 6 7 12 13 14 15 20 21 22 XX Sex chromosomes E.Schmid/2001 Examples of genetic disorders in humans due to changes in chromosome numbers or chromosome structures In the following section we will look at prominent disorders in humans which are caused by chromosomal aberrations and which can be diagnosed with the help of the karyotyping method 1. Trisomy 21 (= Down syndrome) - characterized by cells which show an extra copy of the chromosome 21 in a karyogram - together with Trisomy 13, the most common chromosome number abnormality in humans - affects about 1 out of every 700 children born in the US - genetic basis of the rare human disorder called Down syndrome - affected people suffer from heart defects, susceptibility to respiratory infections, leukemia and usually have a shorter life-span - many affected individuals exhibit varying degrees of mental retardation - incidence of Down syndrome in the offspring of genetically normal parents increases markedly with the age of the mother - the chromosome alterations of Trisomy 21 are suspected to occur after fertilization 33 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 2. Trisomy 13 (Pateu’s syndrome) - Affected human individuals are characterized by cells which show an extra copy of the chromosome 13 in a karyogram (see red circle in Figure below) - This chromosomal aberration is observed in about 1 out of every 5000 life births - Trisomy 13 babies are frequently stillborn or die as newborns - Physical abnormalities include: severe mental retardation, growth retardation, mis-development of the brain/spinal cord, cleft lip and palate, cyclopia (one eye) often with protruding facial proboscis (see Figure below) Karyotype: Trisomy 13 Examples of clinical features of Trisomy 13: 34 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 3. Trisomy 18 (= Edwards syndrome) - characterized by cells which show an extra copy of the chromosome 18 in a karyogram - 2nd most common autosomal aberration; incidence: 1/8000 live births - severe facial (cleft lip, microcephalus) and skeletal malformations - many organ malfunctions - 90% of children with Trisomy 18 die by 12 months of age in most cases of severe chromosomal aberrations or abnormal chromosomal numbers, e.g. Triploidies, however, the affected human embryo is spontaneously aborted long before birth (= miscarriage) all Trisomies, except Trisomie 21, produce infants who will usually die during the first few months of life around half of all early spontaneous abortions are due to Trisomies 4. Fragile X-syndrome (Martin-Bell syndrome) - The most common form of inherited mental retardation - Affected individuals have cognitive/behavioral/neuropsychological problems mild-to-moderate autistic-like behavior - occurs in 1/1500 male and 1/2500 female life births - clinical phenotype results from Fragile X-chromosomal ends (Xq27.3) - A short, extra chromosomal fragment is observable at one of the two long sister chromatid arms of the X-chromosome - This “chromosomal appendix” is caused by excessively long DNA inserts, so-called (CGG) n repeats in the coding sequence of the FMR-1 gene which is located at the X-chromosomal end normal: pre-mutated fully mutated - 5-55 CGG repeats 65-230 CGG repeats > 230 CGG repeats the karyotype is characterized by a thin strand of genetic material which extends beyond the long arm at the highly conserved Xq27.3bof the Xchromosome makes the X chromosome fragile looking 5. Ataxia telangietactica (AT) or Louis-Bar syndrome - autosomal recessive inherited disorder - characterized by chromosomal defects, most importanlty translocations affecting the chromsomes # 7 and # 14 - juvenile development of excessive blood vessels (= telangiectasias) in the conjuctivae of the eyes - immune defiency leads to heavy, recurrent infections of the lungs and the bronchial system - many individuals suffering from AT have a high tendency (predisposition) to develop tumors (lymphomas, leukemias) - AT patients are extremely sensitive to X-rays and strong irradiation 35 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 6. Chromosome 5p deletion syndrome or (“cat cry – cri du chat syndrome”) - cri du chat syndrome is caused by a deletion of the short (= p) arm of the human chromosome 5 - affects 1/20,000 to 1/50,000 human life births - infants with cri du chat syndrome commonly have a distinctive cat-like cry - show severe mental retardation - low birth weight, microcephaly, webbed fingers or toes - cause is not known spontaneous loss of a piece of chromosome 5 during development of an egg or sperm 7. Chromosome 13q deletion - relatively common finding in human blood cancers, such as: - B-cell chronic lymphocytic leukemia (CLL), non-Hodgkin's lymphoma (NHL) and multiple myeloma (MM) Image of chromosome #13 deletion 8. Chromosome 4p deletion (= Wolf-Hirschhorn syndrome) - human disorder first described by Wolf (Freiburg, Germany) and Hirschhorn (New York, U.S.A.) - caused by mostly “de novo” deletion of a tiny chromosomal fragment from the short arm of chromosome # 4 - affected children show pronounced physical and psychological impairments with retarded development - body shows several deformations affecting the face, but also scalp and heart defects may be observed 9. 14/21 Translocation carrier - most common translocation leading to Down syndrome - caused by an exchange = translocation between chromosome #14 and #21 - almost the complete chromosome #21 is attached to the short (p) arm of chromosome #14 - affected individuals are phenotypic normal but bear high risk of having a Down syndrome child 36 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 10. 22q-“Philadelphia” chromosome translocations - chromosomal anomalies which lead to certain forms of blood cancers, e.g. acute lymphatic leukemia (ALL) - it is caused by translocation of a fragment of the long arm of chromosome 22 over to chromosome 9 - the t(9;22) translocation event brings two genes called c-ABL (chr. #9q34) and BCR (chr. # 22q11) together and forms a new hybrid gene - the expressed hybrid gene leads to a 190 or 210 kDa so-called fusion protein Bcr-Abl, which plays a role in cancer development Formation of the “Philadelphia chromosome” by translocation non-disjunction in meiosis does not only affect autosomes , such as chromosome 21, but can also lead to abnormal numbers of sex chromosomes this has usually less dramatic effects on the genetic balance and on the phenotype of the carrier than unusual numbers of autosomes this probably due to the lesser number of genes on sex chromosomes and the natural inactivation of the second X-chromosome in an XX (= female) individual the most common sex chromosome abnormalities are: 1. Extra Y chromosome (XYY) - observed in 1 out of 2500 live births - affected males have larger body stature and borderline intelligence - new studies give hints to mild to severe social behavioral disturbances with trend to accumulated criminal records 37 SAN DIEGO MESA COLLEGE SCHOOL OF NATURAL SCIENCES Introduction to Molecular & Cell Biology; Instructor: Elmar Schmid, Ph.D. 2. XXY = Klinefelter syndrome (XXXY or XXYY) - cells of human males show an extra X chromosome - this chromosomal aberration is observed in 1 out of 2000 live births - affected males have abnormally small testes and the affected individual is sterile - often accompanied by breast enlargement Karyotype of a human individual with Klinefelter syndrome X X Y 3. XXX = Multi-X female (“super-female”) - carriers have limited fertility - around one woman in 1000 has three or more X-chromosomes - most 47,XXX women are ‘normal’ - the 48,XXXX woman are usually mildly retarded, and 49,XXXXX produces severe disability 4. XO = Turner syndrome - affected individuals are females which show underdeveloped ovaries, poor breast development and a so-called web of skin between neck and shoulders - carriers are sterile 38