chapter12 summary
advertisement

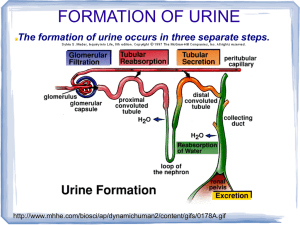
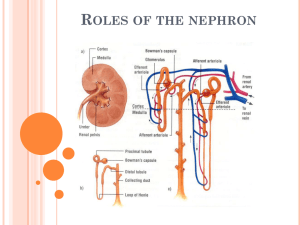
CHAPTER12 SUMMARY •Excretion is crucial to internal fluid homeostasis in the following ways: (1) Maintenance of proper internal levels of inorganic solutes (Na +, K+, Cl-, H+, CO2, etc.); (2) Maintenance of proper water volume in body fluids, important for proper circulatory-fluid pressure and cell hydration; (3) removal of non-nutritive and harmful substances resulting from metabolism or ingestion without losing useful organic molecules; (4) maintenance of osmotic balance (which results from a combination of water and dissolved solutes). Four types of organ systems have evolved to regulate excretion: •Respiratory systems (gills and lungs) regulate CO2. Gills can remove other solutes such as ammonia, and can help regulate HCO3-. •Digestive systems remove not only undigested food but also some end-products of internal metabolism such as bilirubin from the vertebrate liver. Intestinal tracts may also help regulate ions and water. •Integument (skin) and glands may excrete some organic wastes as secondary functions (such as sweat glands in humans), or as primary functions to excrete excess inorganic ions (salt glands of brine shrimp and marine birds and reptiles). •Renal organs (e.g., crustacean antennal glands, insect Malpighian tubules, and vertebrate kidneys) have tubules that filter body fluids and regulate water, ions and many organic substances, and then selectively reabsorb or secrete these molecules. The output of renal organs is usually called urine. •Nitrogen metabolism creates special stresses and produces three major end products: •Ammonia is the primary nitrogenous product of protein breakdown. It is highly toxic and requires considerable water to flush out. Thus it is the primary nitrogenous waste only in most aquatic animals •Urea (formed from two ammonias and one carbon dioxide) costs energy to make, but is less toxic and requires less water to flush out. It is the primary nitrogenous waste in mammals, adult amphibians, and some reptiles. It is also a major osmolyte in cartilaginous fishes. •Uric acid (formed from purine metabolism, containing 4 nitrogens) is the most costly nitrogenous waste, but is non-toxic and may be beneficial as an antioxidant. It is the primary nitrogenous waste in birds, most reptiles, and insects. •Renal excretory organs use up to four processes to produce urine: (1) Filtration, in which water and small solutes pass through a sieve-like boundary. Filtration can be driven by either hydrostatic or osmotic pressure differences across the boundary. Hydrostatically driven filtration is also called ultrafiltration; (2) Secretion, in which specific solutes are transported into the tubule lumen for excretion; (3), in which specific solutes and water may be transported back into the body from the lumen; (4) Osmoconcentration, in which water is removed from the lumen fluid while leaving solutes behind, producing an excretion more concentrated than body fluids and thus saving water. Tubule functions are studied in isolated perfused tubules. Types are: •Protonephridia (in rotifers, flatworms, larval annelids and larval mollusks), which use ultrafiltration driven by cilia, secretion, and reabsorption •Metanephridia (in adult mollusks, crustaceans, and vertebrates), which use ultrafiltration typically driven by blood pressure, secretion, reabsorption, and sometimes osmoconcentration. The mammalian kidney is a methanephridial organ. •Malpighian tubules and hindguts (in terrestrial arthropods), which use filtration driven by osmosis, secretion, reabsorption, and sometimes osmoconcentration Mammalian Urinary System The urine-forming functional unit of the kidneys is the nephron, which is composed of interrelated vascular and tubular components. •The vascular component consists of two capillary networks in series, the first being the glomerulus, a tuft of capillaries that filters large volumes of protein-free plasma into the tubular component. •The second capillary network consists of the peritubular capillaries, which wind around the tubular component. The peritubular capillaries nourish the renal tissue and participate in exchanges between the tubular fluid and plasma. •The tubular component begins with Bowman’s capsule, which cups around the glomerulus to catch the filtrate, then continues a specific tortuous course to ultimately empty into the renal pelvis. As the filtrate passes through the various regions of the tubule, the cells lining the tubules modify it, adding some unwanted ECF constituents, and returning to the plasma only those materials necessary for maintaining the proper extracellular fluid composition and volume. What is left behind in the tubules is excreted as urine. •The kidneys perform four basic processes in carrying out their regulatory and excretory functions: (1) glomerular filtration, the nondiscriminating movement of protein-free plasma from the blood into the tubules; (2) tubular reabsorption, the selective transfer of specific constituents in the filtrate back into the blood of the peritubular capillaries; (3) tubular secretion, the highly specific movement of selected substances from the peritubular capillary blood into the tubular fluid; and (4) osmoconcentration, producing a concentrated extracellular fluid and urine. Everything that is filtered or secreted but not reabsorbed is excreted as urine. Glomerular Filtration •Glomerular filtrate is produced as a portion of the plasma flowing through each glomerulus is passively forced under pressure through the glomerular membrane into the lumen of the underlying Bowman’s capsule. The net filtration pressure that induces filtration is caused by an imbalance in the physical forces acting across the glomerular membrane. A high glomerular capillary blood pressure favoring filtration outweighs the combined opposing forces of plasmacolloid osmotic pressure and Bowman’s capsule hydrostatic pressure. •Of the plasma flowing through the kidneys, normally 20% is filtered through the glomeruli, producing an average glomerular filtration rate (GFR) of 125 ml/min in humans. This filtrate is identical in composition to plasma except for the plasma proteins retained by the glomerular membrane. •The GFR can be deliberately altered by changing the glomerular capillary blood pressure as a result of sympathetic influence on the afferent arterioles. Afferent arteriolar vasoconstriction decreases the flow of blood into the glomerulus, resulting in a reduction in glomerular blood pressure and a fall in the GFR. Conversely, afferent arteriolar vasodilation leads to increased glomerular blood flow and a rise in the GFR. Sympathetic control of the GFR is part of the baroreceptor reflex response to compensate for a change in arterial blood pressure. As the GFR is altered, the amount of fluid lost in the urine is changed correspondingly, providing a mechanism to adjust plasma volume as needed to help restore blood pressure to normal on a longterm basis. Tubular Reabsorption •After a protein-free plasma is filtered through the glomerulus, each substance is handled discretely by the tubules; the concentrations of different constituents are variously altered as the filtered fluid flows through the tubular system. •The reabsorptive capacity of the tubular system is tremendous. Over 99% of the filtered plasma is returned to the blood through reabsorption. •The major substances actively reabsorbed are Na+ (the principal extracellular fluid cation), most other electrolytes, and organic nutrients such as glucose and amino acids. The most important passively reabsorbed substances are Cl- H2O, HCO3-, and urea. •The pivotal event to which most reabsorptive processes are linked in some way is the active reabsorption of Na+. An energy-dependent Na+–K+ ATPase carrier located in the basolateral membrane of each proximal tubular cell transports Na+ out of the cells into the lateral spaces between adjacent cells. This transport of Na+ induces the net reabsorption of Na + from the tubular lumen to the peritubular capillary plasma, most of which takes place in the proximal tubules. The energy used to supply the Na+–K+ ATPase carrier is ultimately responsible for the reabsorption from the proximal tubule of Na+ glucose, amino acids, Cl-, HCO3-, H2O, and urea. •Specific cotransport carriers located at the luminal border of the proximal tubular cell are driven by the Na+ concentration gradient to selectively transport glucose or an amino acid from the luminal fluid into the tubular cell, from which the nutrient eventually enters the plasma. •Chloride is passively reabsorbed down the electrical gradient established by active Na + reabsorption. •Water is passively reabsorbed as a result of the osmotic gradient created by active Na+ reabsorption. Sixty-five percent of the filtered H2O is reabsorbed from the proximal tubule in this unregulated fashion. This extensive reabsorption of H 2O increases the concentration of other substances remaining in the tubular fluid, most of which are filtered waste products. •About 50% of filtered urea (a waste product of protein metabolism) is reabsorbed. The other waste products, failing to be reabsorbed, remain in the urine in highly concentrated form. •The other electrolytes actively reabsorbed by the tubules, such as PO 43_ and Ca++, have their own independently functioning carrier systems. Once the filtered load of an actively reabsorbed substance exceeds the transport capacity ( Tm), reabsorption proceeds at a constant maximal rate, with the additional filtered quantity of the substance being excreted in the urine. •Na+ reabsorption is regulated in the proximal and distal tubules and collecting ducts, though less than half the filtered Na+ is regulatable. The extent of this controlled Na+ reabsorption depends on the renin-angiotensin-aldosterone system. Because Na + and its attendant anion, Cl-, are the major osmotically active ions in the extracellular fluid (ECF), the ECF volume is determined by the Na+ load in the body. In turn, the plasma volume, which reflects the total ECF volume, is important in the long-term determination of arterial blood pressure. Whenever the Na+ load/ECF volume/plasma volume/arterial blood pressure are below normal, the kidneys secrete renin, an enzymatic hormone that triggers a series of events ultimately leading to increased secretion of aldosterone from the adrenal cortex. Aldosterone increases Na reabsorption from the proximal and distal tubules, correcting for the reduction in Na/ECF volume/blood pressure. Tubular Secretion •By means of the process of tubular secretion, the kidney tubules are able to selectively add some substances to the quantity already filtered. Secretion of substances hastens their excretion in the urine. •The most important secretory systems are for (1) H +, which is important in the regulation of acid-base balance; (2) K+, which keeps the plasma K+ concentration at an appropriate level to maintain normal membrane excitability in muscles and nerves; and (3) organic ions, which accomplishes more efficient elimination of foreign organic compounds from the body. •Only a small fraction of filtered fluid remains in the tubules to be excreted as urine. Only wastes and excess electrolytes not wanted by the body are left behind, to be eliminated in the urine. Because the excreted material is “cleared” from the plasma, the term plasma clearance refers to the volume of plasma being cleared of a particular substance each minute by means of renal activity. Osmoconcentration •The kidneys are able to excrete urine of varying volumes and concentrations to either conserve or eliminate H2O, depending on whether the body has a H2O deficit or excess, respectively. Human kidneys are able to produce urine ranging from 0.3 ml/min at 1,200 mOsm to 25 ml/min at 100 mOsm by reabsorbing variable amounts of H2O from the distal portions of the nephron. •This variable reabsorption is made possible by the establishment of a vertical osmotic gradient ranging from 300 to 1,200 mOsm (in humans) in the medullary interstitial fluid by means of the loop of Henle countercurrent system and urea recycling between the collecting duct and Henle’s loops. This vertical osmotic gradient to which the hypotonic (100 mOsm) tubular fluid is exposed as it passes through the distal portions of the nephron establishes a passive driving force for progressive reabsorption of H2O from the tubular fluid, but the actual extent of H2O reabsorption depends on the amount of vasopressin (antidiuretic hormone) secreted. •Vasopressin increases the permeability of the distal tubules and collecting ducts to H 2O; they are impermeable to H2O in its absence. Vasopressin secretion increases in response to a H 2O deficit, and H2O reabsorption increases accordingly. Vasopressin secretion is inhibited in response to a H2O excess, thereby reducing H2O reabsorption. In this way, adjustments in vasopressincontrolled H2O reabsorption help correct any fluid imbalances. •Different osmoconcentrating abilities among species depend on nephron anatomy and metabolic rates. Desert rodents, for example, have relatively long nephron loops and high metabolic rates, and can concentrate urine up to 6,000 mOsm. Peristaltic pumping by the renal pelvis may contribute to the concentrating process. Bladder Storage and Micturition •Once formed, urine is propelled by peristaltic contractions through the ureters from the kidneys to the urinary bladder for temporary storage. The bladder can accommodate a relatively large volume of urine before stretch receptors within its wall initiate the micturition reflex. This reflex causes involuntary emptying of the bladder by simultaneous bladder contraction and opening of both the internal and external urethral sphincters. Micturition can transiently be voluntarily prevented until a more opportune time for bladder evacuation by deliberate tightening of the external sphincter and surrounding pelvic diaphragm. Other Vertebrate Urinary Systems and Extrarenal Organs •Freshwater bony fishes must maintain an internal osmotic pressure far above that of the environment, and are faced with constant influx of water through the gills and mouth. Their kidneys excrete a highly dilute voluminous urine. Dietary intake and active transport by the gills brings in NaCl and other ions to replace those lost in the urine. Thus the gills serve as extrarenal osmoregulatory organs. •Marine bony fishes use their gills for most excretory functions. Most are highly hypoosmotic compared to seawater, so they face constant influx of excess salt through the gills and diet, and constant water loss through the gills. Gills compensate for this, using specialized epithelial chloride cells, which actively transport NaCl outwards. •Cartilaginous fishes are isosmotic or slightly hyperosmotic relative to seawater, because they retain urea and TMAO as major osmolytes (Chap. 13). The gills are relatively impermeable to urea and TMAO, and the nephrons reabsorb most of these osmolytes from the filtrate, excreting only what is necessary for nitrogen balance. Extrerenal organs perform most other excretion and osmoregulatory roles. Gills are thought to remove some of the excess NaCl, but in particular, these fish have a specialized hindgut organ called the rectal gland, which excrete a hypertonic fluid high in NaCl. •Amphibians have kidney nephrons similar to those of freshwater fish, with similar functions (to excrete water and reabsorb ions and nutrients). They also excrete urea by filtration and tubular secretion. In terrestrial amphibians, the urinary bladder has a secondary role, that of a temporary water reservoir. With dehydration stress, the release of the hypothalamic hormone arginine vasotocin (AVT) triggers water uptake into the main body from the wall of the bladder. Uptake through the skin may also occur. •Reptiles have kidney nephrons similar to aquatic vertebrates. The ureters carry urine in a liquid or semisolid form into the cloaca, the final chamber of the hindgut. They can conserve water in three ways (1) uric acid as the primary nitrogenous waste conserves water; (2) the cloaca or lower intestine can reabsorb some water by first transporting salt. This precipitates uric acid even further; (3), marine and some desert reptiles have an extrarenal organ, the nasal salt gland that is dedicated to excreting a highly salty fluid. •Birds have kidneys that send a liquid or semisolid urine into a cloaca, with a function similar to that of reptiles. The kidneys are usually dominated by the typical non-mammalian nephron, called “reptilian type.” However they also have some nephrons, called “mammalian type,” with countercurrent loops that can form a concentrated urine. Their primary nitrogenous waste is uric acid. precipitated as crystals which exert no osmotic pressure. Marine birds also have a nasal salt gland located near the eyes, with ducts leading to the nasal passages. These glands transport NaCl out of the blood without concomitant osmotic water movement. Insect Malpighian Tubules •These blind-end ducts in terrestrial arthropods (e.g., insects) project into the hemolymph from the hindgut. They filter the hemolymph by secreting ions into the lumen. This makes the tubule fluid more concentrated, so water from the hemolymph moves in by osmosis. •Wastes such as uric acid are also transported into the lumen. The tubule empties into the hindgut, where epithelial cells of the rectum can further modify the urine, often osmoconcentrating it (i.e., making it hyperosmotic to conserve water). Uric acid precipitates as crystals as water is removed.