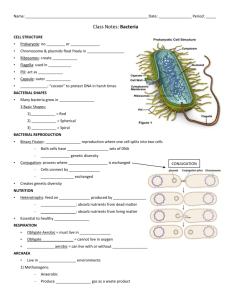
SIGNAL SEQUENCE DIRECTS LOCALIZED SECRETION OF
BACTERIAL SURFACE PROTEINS
Fredric Carlsson, Margaretha Stålhammar-Carlemalm, Klas Flärdh, Charlotta Sandin, Eric
Carlemalm and Gunnar Lindahl
SUPPLEMENTARY METHODS
Bacterial strains and culture conditions. E. coli strains XL-1 and KJ622 were used for
subcloning. Strain KJ622 significantly lowers the copy number of plasmids replicating via the
ColE1 origin, and was used to clone constructs that appeared toxic for XL-11(ref. 1). The
ColE1-derived vector pBR322 and the E. coli-S. pyogenes shuttle vector pLZ12Spec2(ref. 2)
were used for cloning. E. coli was grown in LB broth, with antibiotics added as appropriate. S.
pyogenes was grown in Todd-Hewitt broth supplemented with 0.2% yeast extract (THY),
except in the analysis shown in Fig 4S, which employed C-medium3.
Construction of new S. pyogenes strains. For analysis of the role of signal sequences in
localized secretion, four strains were constructed, in which either of the parental proteins M6
and PrtF, or a domain swap derived from these proteins, was expressed from plasmid
pLZ12Spec, in S. pyogenes strain SAM2. The genes for the wild type proteins were amplified
by PCR from strain JRS4 and ligated into pLZ12Spec, which was transformed into SAM2. For
construction of the domain swap FM, in which the signal sequence of PrtF is fused to the
mature part of M6, a PCR product corresponding to the mature part of M6, except the six most
N-terminal amino acids, was cloned into pLZ12Spec and transformed into E. coli XL-1.
Subsequently, a PCR fragment encoding the signal sequence of PrtF, and four amino acids in
PrtF located C-terminally of the signal peptidase cleavage site, was cloned in frame with the
fragment encoding the mature part of M6. This PCR fragment also included the promoter
1
region of the gene encoding PrtF. The four amino acids derived from the mature part of PrtF
were retained to keep intact the signal peptidase recognition site. To compensate for these four
residues, and for a sequence of two additional amino acids (LE) encoded by an XhoI site
introduced during the cloning work, the six most N-terminal amino acids were excluded from
the mature part M6. The structure of the gene encoding this domain swap was verified by DNA
sequencing. Finally the construct was transformed into SAM2 and a clone, designated
SAM2/pFM, was identified by PCR and by analysis of protein expression. The reciprocal
domain swap, designated MF, in which the signal sequence of M6 is fused to the mature part
of PrtF, was constructed by a similar strategy. However, due to toxicity problems in E. coli,
plasmid pBR322 and strain KJ622 were used for gene construction work, and 10 amino acid
residues were kept C-terminally to the signal peptidase cleavage site in M6. A correct construct
was transferred from pBR322 to pLZ12Spec and the resulting plasmid was transformed into E.
coli KJ622. After verification of the gene structure by DNA sequencing, this plasmid was
transformed into S. pyogenes SAM2, and a transformant, designated SAM2/pMF, was
identified as described above.
The role of the conserved signal sequence motif YSLRK in M protein secretion was
analyzed with the M5 protein4. Of note, the sequence of this motif in M5 was initially4
reported to be YWLRK, but our repeated analysis of the chromosomal emm5 gene showed that
the correct sequence is YSLRK, as in many other M proteins. Two mutant strains were used, in
which the motif was changed from YSLRK to YAARK or YSAAK. Thus, two amino acid
residues were changed in each mutant. For construction of the mutants, plasmid pLZ12Spec
carrying the emm5 gene5 was subjected to site-specific mutagenesis using the QuickChange kit
(Stratagene), and the plasmid carrying the wild type gene and the two mutant plasmids were
used to transform the M-negative strain M5 (ref. 5). The three plasmid-carrying strains are
referred to as pM5 YSLRK, pM5 YAARK and pM5 YSAAK.
2
Purified proteins and antisera. Human Fn, a 30 kDa Fn-fragment, trypsin (from bovine
pancreas), and gold-labeled staphylococcal protein A (20 nm colloidal gold) were from Sigma.
Unlabeled protein A and gold-labeled goat anti-rabbit IgG (15 nm colloidal gold) were from
Amersham. Purified M5 protein6 was used to raise rabbit antiserum as described7. Because of
the extensive residue identity between the M5 and M6 proteins this antiserum also recognizes
M6, and it is referred to here as anti-M. Rabbit anti-human Fn was from DakoCytomation.
Rabbit antiserum against Bacillus subtilis SecA was the kind gift of Dr. R. Freudl. This antiSecA serum, which had been raised against highly purified protein8, did not react with
streptococcal M protein (data not shown).
Binding assays with bacteria. Surface-localized M protein was detected by incubation of
whole washed bacteria with rabbit anti M-serum, as described9, using 125I-labeled protein A to
detect bound rabbit antibodies. Binding of
125
I-labeled Fn (30 kDa fragment) to whole
streptococci was analyzed as described10. Of note, PrtF is the only Fn-binding protein
expressed by the S. pyogenes M6 strain, allowing the use of Fn for detection of surfacelocalized PrtF11(ref. 11 and our unpublished data). Binding is expressed as percent of added
radioactivity. The data in Fig. 2c and in Supplementary Fig. 3b are based on three different
experiments with duplicate samples and are presented as means ± SD.
Extraction of surface proteins from S. pyogenes. The M6 protein expressed by strain
SAM2/pFM was extracted by pepsin digestion at suboptimal pH12(ref. 12). This method of
preparation explains the low MW of the protein fragment recovered. The PrtF expressed by S.
pyogenes strains JRS145 and SAM2/pMF was extracted by a technique developed for this
purpose, using 8 M urea. In brief, a bacterial culture (4500 ml) in THY was grown to A620 ≈
0.9, the bacteria were washed twice with 50 mM Tris-HCl, pH 7.3, resuspended to 2 x 1010
bacteria/ml in the same buffer supplemented with 8 M urea, and rotated at room temperature
for 2 h. After centrifugation, the supernatant was collected and dialyzed against PBS. For
3
partial purification of PrtF, the extract was applied to a 1 ml HiTrap column (Amersham
Pharmacia) containing immobilized Fn. Bound proteins were eluted with 3 M KSCN and the
eluate was dialyzed against PBS. Proteins were identified by SDS-PAGE and Western blot.
The amino-terminal sequence of proteins on blotting membranes was determined at the Protein
Analysis Centre of Karolinska Institutet (Stockholm, Sweden).
Detection of surface proteins by immunofluorescence. A culture (10 ml) of the S. pyogenes
strain indicated was grown to late logarithmic phase (A620 ≈ 0.8). The bacteria were washed
once with PBS containing 0.025% Tween-20 (PBST), resuspended in PBST (1 ml) with or
without trypsin (0.2 mg/ml) and incubated at 37C for 30 min. Samples of the untreated and
the trypsinized bacteria were immediately taken for analysis, and the remaining trypsinized
bacteria were washed and reinoculated into pre-warmed THY (10 ml) and incubated at 37C
for 15 min, to allow protein synthesis. All three bacterial samples were washed twice with
PBST containing 0.02% azide (PBSAT), resuspended in PBSAT to 2 x 10 9 bacteria/ml, and
used for detection of surface proteins. For this analysis, a sample (100 l) of the washed
bacteria was incubated for 1 h at room temperature with 10 µl rabbit anti-M serum (final
dilution 1/100) or 10 µl of a solution containing the 30 kDa Fn fragment (500 µg/ml). The
bacteria incubated with the Fn fragment were subjected to a second incubation with anti-Fn
antibodies; for this purpose washed bacteria were resuspended in PBSAT (100 µl) containing
rabbit anti-human Fn (final dilution 1/20) and the suspension was incubated for 1 h at room
temperature. The samples incubated with anti-M serum or anti-Fn serum were washed and
bound rabbit antibodies were detected with Alexa fluor 488-conjugated goat anti-rabbit IgG
(Molecular Probes). Fading of the fluorochrome was reduced by the use of SlowFade® light
antifade kit (Molecular Probes). Images were obtained using a Zeiss AxioImager.Z1
microscope (Carl Zeiss), appropriate filters, a Photometrics CoolSNAP fx CCD camera, and
4
Zeiss AxioVision 4 software. Adobe Illustrator version 10 was used for final figure
preparation.
Immunogold electron microscopy. Bacteria were prepared as described for IF, using rabbit
antibodies to detect surface-localized M6 and PrtF. Bound antibodies were detected by
incubation with gold-labelled protein A (final dilution 1/15). After incubation for 1 h at room
temperature, the bacteria were washed once with PBSAT and once with PBSAT that had been
diluted two-fold in ddH2O. After this incubation, the bacteria were adsorbed to carbon film on
400 mesh copper grids and observed at 100 kV in a Philips CM 10 transmission electron
microscope without further added contrast. To avoid bias, images were taken by a person not
knowing the source of the preparation analyzed. In the electron micrographs, contrast is
inverted to improve visualization of the new septum.
Analysis of cellular distribution of SecA. The specificity of the anti-SecA serum used was
analyzed with a whole-cell lysate of S. pyogenes. For this purpose, a lysate was prepared from
JRS4 bacteria (grown to late logarithmic phase) by treatment with mutanolysin (0.4 mg/ml;
Sigma) in the presence of a protease inhibitor cocktail (Sigma). A sample was boiled in sample
buffer and analyzed by Western blot with anti-SecA. For the EM analysis shown in Fig. 4b,
bacteria were grown in THY to late logarithmic phase (A620 ≈ 0.8). For the EM analysis shown
in Supplementary Fig. 4S, bacteria were grown to early stationary phase, when the SpeB
protein is expressed13, using C medium, which was used for identification of the Exportal3. The
bacteria were washed in PBST and fixed in a mixture of 1.5% glutaraldehyde and 1.5%
formaldehyde in 0.1 M phosphate buffer pH 7 for 1 h and embedded in Lowicryl HM2014.
Sections were cut on a LKB SuperNova ultramicrotome and collected on pioloform coated 400
mesh grids. Rabbit anti-SecA antiserum and gold-labeled goat anti-rabbit IgG were used for
detection of SecA. The sections were post stained with 4% uranylacetate and observed at 60
5
kV in a Philips CM 10 Microscope. Adobe Illustrator version 10 was used for figure
preparation.
6
Supplementary references
1. Perez, A. R., Abanes-De Mello, A. & Pogliano, K. SpoIIB localizes to active sites of septal
biogenesis and spatially regulates septal thinning during engulfment in Bacillus subtilis. J.
Bacteriol. 182, 1096-108 (2000).
2. Husmann, L. K., Scott, J. R., Lindahl, G. & Stenberg, L. Expression of the Arp protein, a member
of the M protein family, is not sufficient to inhibit phagocytosis of Streptococcus pyogenes. Infect.
Immun. 63, 345-8 (1995).
3. Rosch, J. & Caparon, M. A microdomain for protein secretion in Gram-positive bacteria. Science
304, 1513-5 (2004).
4. Miller, L., Gray, L., Beachey, E. & Kehoe, M. Antigenic variation among group A streptococcal M
proteins. Nucleotide sequence of the serotype 5 M protein gene and its relationship with genes
encoding types 6 and 24 M proteins. J. Biol. Chem. 263, 5668-73 (1988).
5. Johnsson, E. et al. Role of the hypervariable region in streptococcal M proteins: binding of a human
complement inhibitor. J. Immunol. 161, 4894-901 (1998).
6. Johnsson, E. et al. A highly variable region in members of the streptococcal M protein family binds
the human complement regulator C4BP. J. Immunol. 157, 3021-9 (1996).
7. Berggård, K. et al. Binding of human C4BP to the hypervariable region of M protein: a molecular
mechanism of phagocytosis resistance in Streptococcus pyogenes. Mol. Microbiol. 42, 539-51
(2001).
8. Klose, M., Schimz, K. L., van der Wolk, J., Driessen, A. J. & Freudl, R. Lysine 106 of the putative
catalytic ATP-binding site of the Bacillus subtilis SecA protein is required for functional
complementation of Escherichia coli secA mutants in vivo. J. Biol. Chem. 268, 4504-10 (1993).
9. Stålhammar-Carlemalm, M., Stenberg, L. & Lindahl, G. Protein Rib: a novel group B streptococcal
cell surface protein that confers protective immunity and is expressed by most strains causing
invasive infections. J.Exp. Med. 177, 1593-603 (1993).
7
10. Carlsson, F., Berggård, K., Stålhammar-Carlemalm, M. & Lindahl, G. Evasion of phagocytosis
through cooperation between two ligand-binding regions in Streptococcus pyogenes M protein. J.
Exp. Med. 198, 1057-68 (2003).
11. Hanski, E. & Caparon, M. Protein F, a fibronectin-binding protein, is an adhesin of the group A
streptococcus Streptococcus pyogenes. Proc. Natl Acad. Sci. USA 89, 6172-6 (1992).
12. Beachey, E. H. et al. Purification and properties of M protein extracted from group A streptococci
with pepsin: covalent structure of the amino terminal region of type 24 M antigen. J. Exp. Med. 145,
1469-83 (1977).
13. Lyon, W. R. & Caparon, M. G. Trigger factor-mediated prolyl isomerization influences maturation
of the Streptococcus pyogenes cysteine protease. J. Bacteriol. 185, 3661-7 (2003).
14. Carlemalm, E., Garavito, R. & Villiger, W. Resin development for electron microscopy and an
analysis of embedding at low temperature. J. Microsc. 126, 1213-143 (1982).
8
 0
0