ele12529-sup-0003-AppendixS3
advertisement

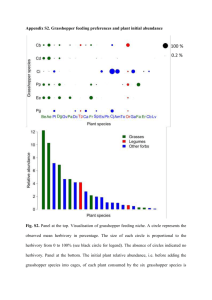
Appendix S3. Herbivore survival METHODS Grasshopper survival in intra and inter-specific treatments Using survivorship data, we compared the survival of each grasshopper species in monospecific treatments to its survival in the 3- and 6- species treatments to quantify species performance in inter-specific treatment relatively to intra-. Grasshopper response to inter- vs. intra-specific treatments were quantified by calculating the log response ratio for each grasshopper species following Suding et al. (2003): 𝐺𝑟𝑎𝑠𝑠ℎ𝑜𝑝𝑝𝑒𝑟𝑠𝑝𝑒𝑐𝑖𝑒𝑠𝑠𝑢𝑟𝑣𝑖𝑣𝑎𝑙𝑖𝑛𝑚𝑢𝑙𝑡𝑖𝑠𝑝𝑒𝑐𝑖𝑒𝑠𝑡𝑟𝑒𝑎𝑡𝑚𝑒𝑛𝑡 𝐿𝑁𝑅𝑅𝑠𝑢𝑟𝑣𝑖𝑣𝑎𝑙 = ln(𝐺𝑟𝑎𝑠𝑠ℎ𝑜𝑝𝑝𝑒𝑟𝑠𝑝𝑒𝑐𝑖𝑒𝑠𝑠𝑢𝑟𝑣𝑖𝑣𝑎𝑙𝑖𝑛𝑚𝑜𝑛𝑜𝑠𝑝𝑒𝑐𝑖𝑓𝑖𝑐𝑡𝑟𝑒𝑎𝑡𝑚𝑒𝑛𝑡) A LNRRsurvival > 0 indicates that grasshopper species survival was higher when interacting with other species than when interacting with itself, a pattern that can emerge if feeding niche complementarity occurs between grasshoppers (Deraison et al. 2015). A LNRRsurvival < 0 indicates the opposite. Complementarity between grasshoppers is predicted to be higher when grasshopper exhibited contrasted set of traits, assuming that trait differences between species reflect species niche differences (Maire et al. 2012) (Fig. 1). We calculated a functional dissimilarity index to test the effect of grasshopper trait dissimilarity on their survival. The index was calculated in each multispecies treatments (3- and 6- species treatments) and for each grasshopper trait (body size, incisor strength and C:N ratio), such as: 𝐷𝑖𝑠 − 𝑇𝑟𝑎𝑖𝑡 = ∑𝑛 𝑗 (𝑇𝑟𝑎𝑖𝑡𝑣𝑎𝑙𝑢𝑒𝑜𝑓𝑡ℎ𝑒𝑠𝑝𝑒𝑐𝑖𝑒𝑠𝑖− 𝑇𝑟𝑎𝑖𝑡𝑣𝑎𝑙𝑢𝑒𝑜𝑓𝑡ℎ𝑒𝑠𝑝𝑒𝑐𝑖𝑒𝑠𝑗 )² 𝑁𝑢𝑚𝑏𝑒𝑟𝑜𝑓𝑝𝑎𝑖𝑟𝑤𝑖𝑠𝑒𝑠𝑝𝑒𝑐𝑖𝑒𝑠 Dis-Trait is thus the average difference in trait value of a grasshopper species i from its competing species j divided by the number of pairwise interactions in a given grasshopper mixture. We used three models to assess the relative performance of grasshopper species in inter- vs. intra-specific treatments (LNRRsurvival). We first tested whether the six different grasshopper species showed contrasting survival in inter-specific treatments compared to intra- as a function of their species identity and sex. The second model tested the effect of the mean trait value of the sex/species (body size, incisor strength and C:N ratio) and trait dissimilarity (Dis-BodySize, Dis-IncisorStrength and Dis-C:N ratio). The third model included the effect of plant community functional structure (mean plant height, mean plant C:N ratio, and plant functional diversity) assuming that interactions between grasshoppers are modulated by the local resources (Ritchie & Tilman 1993). To model LNRRsurvival we used mixed effect models. Year and block were included as fixed effects (n = 480). Cage number was included as a random factor. All explanatory variables were standardized (mean-centred and divided by the standard deviation) and model residuals were inspected for normality. In all models, interaction terms between year and explanatory variables were included but they were not significant. The same methodology as explained in the main text was used for model selection and parameter estimation. Post hoc analyses were performed as described in the main text. This analysis focuses on the relative strength of inter-specific interactions between grasshoppers relatively to intra- as it is expected, following the complementarity hypothesis, that increasing niche difference between interacting species leads to higher performance in inter-specific treatment relatively to intra-. Note that this pattern can emerge from two distinct mechanisms, competition and facilitation (Gross et al. 2007), with similar effect on species survival and plant community biomass. We acknowledged that our design based on species replacement and constant herbivore density across treatments cannot formally distinguish between the two mechanisms. RESULTS Survival in inter-specific treatments relatively to intra- (LNRRsurvival) ranged from values close to 0 (i.e. survival in inter-specific treatment = survival in intra-, e.g. in case of C. dorsatus males and females) to significantly positive values (i.e. inter-specific survival > intra-) (Fig S3a). It suggested that survival was generally higher when grasshoppers where in inter-specific mixtures. Grasshopper survival response (LNRRsurvival) was best explained by the functional identity and dissimilarity between herbivores in mixtures as well as the local characteristics of plant communities (Fig. S3b & Table S3). We found a negative effect of herbivore incisor strength and C:N ratio on LNRRsurvival. Herbivore community with weak incisor and low C:N ratio showed a lower survival in intra-specific treatments compared to inter- while survival in intra-specific treatment equal survival in inter- for herbivores with strong incisor and high C:N ratio. We also observed a significant and positive effect of herbivore functional dissimilarity (Dis-Trait). The more the herbivore species differed in their incisor strength (Dis-IncisorStrength), the higher LNRRsurviva was, indicating that herbivore survival was improved in inter-specific mixtures as compared to intra-. Opposite effect was found for herbivore C:N ratio dissimilarity (Dis-C:N ratio). Herbivore body size was not selected in any of the models (Table S3). Finally, we also found that herbivores survival was affected by the functional structure of plant communities. LNRRsurvival decreased with increasing in tall and functional diverse plant community (high mean plant height and plant functional diversity). Thus, survival was higher in inter-specific treatment compare to intra- in small and functionally poor vegetation, while survival became similar in inter- and intraspecific treatment as plant communities get taller and/or functionally richer. Other plant traits (e.g. LDMC, C:N ratio) were not selected in the final model (Table S3). Table S3. Results of linear mixed effect models testing the effect of herbivore functional dissimilarity index (Dis-Trait), herbivore species mean trait values, plant community functional identity (community-weighted means) and diversity on grasshopper survival in inter-specific treatment relatively to intra- (LNRRsurvival). Incisor strength and C:N ratio correspond to the mean trait value per species/sex of herbivore incisor strength and carbon nitrogen ratio respectively. The model also included the cage number (from 1 to 70 cages) as a random effect. LNRRsurvival Model parameters d.f. F value P value Year 1 3.03 0.828 Block 4 0.94 0.361 I(Year-Block) 4 6.67 <0.001 Grasshopper species incisor strength 1 16.40 <0.001 Grasshopper species C:N ratio 1 8.96 <0.001 Dis-Incisor strength 1 4.75 0.007 Dis-C:N ratio 1 6.54 0.008 Mean plant height 1 2.64 0.003 Plant functional diversity 1 2.31 0.07 Residual 464 AICc -221.1 Fig. S3a. Average LNRRsurvival over the two years of the experiment for each grasshopper species and sex (M : males and F: females) (Herbivore species identity: F1,5 = 4.18, P < 0.001; Herbivore species identity: Sex: F1,6 = 6.67, P < 0.001; Year: F1,1 = 3.12, P = 0.07; Block: F1,4 = 0.84, P = 0.50; Year:Block: F1,4 = 5.88, P < 0.001). Significant differences (P < 0.05) between herbivore treatments are represented by different letters (a, b, c), calculated by multiple comparisons for parametric linear mixed effect models. Cb refers to Chorthippus biguttulus, Cd: Chorthippus dorsatus, Ci: Calliptamus italicus, Ee: Euchorthippus elegantulus, Pg: Pezotettix giornae, Pp: Pseudochorthippus parallelus. Fig. S3b. Scaled estimates (±SE) for the grasshopper functional dissimilarity index (DisTrait), grasshopper species mean trait values, plant community functional identity (community-weighted means) and diversity on the LNRRsurvival. Grasshopper species incisor strength and C:N ratio corresponded to the mean trait value per species/sex. Red color indicates variables related to herbivore communities and green color indicates plant community attributes. References 1. Deraison, H., Badenhausser, I., Börger, L. & Gross, N. (2015). Herbivore effect traits and their impact on plant community biomass : an experimental test using grasshoppers. Funct. Ecol., 29, 650–661. 2. Gross, N., Suding, K.N., Lavorel, S. & Roumet, C. (2007). Complementarity as a mechanism of coexistence between functional groups of grasses. J. Ecol., 95, 1296–1305. 3. Maire, V., Gross, N., Börger, L., Proulx, R., Wirth, C., Pontes, L. da S., et al. (2012). Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients. New Phytol., 196, 497–509. 4. Ritchie, M.E. & Tilman, D. (1993). Predictions of species interactions from consumerresource theory: experimental tests with grasshoppers and plants. Oecologia, 94, 516–527. 5. Suding, K.N., Goldberg, D.E. & Hartman, K.M. (2003). Relationships among species traits: Separating levels of response and identifying linkages to abundance. Ecology, 84, 1–16.