Implantion I - Division Of Animal Sciences
advertisement

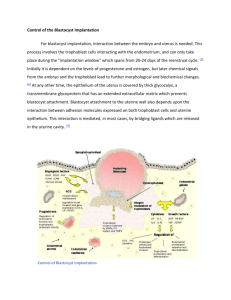
Review Wang and Dey, 2006 Nature Reviews 7:185-199 Lecture X : Comparative Implantation Wang and Dey, 2006 Nature Reviews 7:185-199 Wang and Dey, 2006 Nature Reviews 7:185-199 Implantation Rodents (Mouse) Contact of the blastocyst with the uterus is the first step in the relationship of a mammal embryo with its environment. This contact leads to implantation and placentation. Implantation can be divided into five separate phases: 1. Hatching Phase - Blastocyst loss of Zona pellucida 2. Apposition Phase - Hatched blastocyst aligns and orients itself in preparation for attachment 3. Adhesion Phase - Attachment between uterine surface epithelium and trophoblast 4. Invasion Phase - After blastocyst becomes firmly anchored to the uterine surface, the trophoblast cells begin to penetrate the uterine epithelial layer 5. Postimplantation Phase - Trophoblast cells cease invasive activity and the maternal-fetal relationship is stablized Mesometrial Endometrium Uterine epithelium Zona pellucida ICM Trophectoderm Antmesometrial Blastocyst enters uterus on Day 4 Hatching from the Zona Pellucida Compartive Placentation (ed. D.H. Steven), 1975 Note the closure of the lumen There is extracellular material between uterine epithelium and trophectoderm Apposition Phase Blastocyst comes to lie along the antimesometrial side of uterus. Note that ICM is oriented toward the mesometrial side of uterus. Orientation may be caused by the shape of embryo which is larger at the embryonic pole. Treatment with Relaxin can disrupt the orientation of the blastocysts. Attachment Phase Decidualization begins following adhesion in rodents. Surface changes in the uterine epithelium permit adhesion between trophectoderm and uterine surface epithelium Compartive Placentation (ed. D.H. Steven), 1975 Wang and Dey, 2006 Nature Reviews 7:185-199 Receptivity of Mouse Uterus to Blastocyst Attachment Mucins are large O-linked glycoproteins with ectodomains that extend away from the surface of the cell apical border. Mucins may sterically block access to the cell membrane and function as anti-adhesive molecules. Muc-1 also referred to as epsialin, may function in the time of blastocyst attachment in mammals. Muc-1 is lost from the uterine epithelial apical surface during diestrous in the mouse. It is expressed only on the apical border during the first 3 days of the estrous cycle. Muc-1 expression Muc-1 antibody (green) Perlecan antibody (red) Basement membrane Lumenal Epithelium Glandular Epithelium Muc-1 immunocytochemical staining A. Proestrous B. Estrous C. Metestrous D. Diestrous Only in glandular epithelium, loss of intensity in surface Western blot of Muc-1 Surveyor et al. 1995, Endocrinology 136:3639 Northern Blot A, B, and C = Days 1, 2 and 3 pregnancy D = Day 4 Note: Loss of Muc-1 is associated with time of implantation. Surveyor et al. 1995, Endocrinology 136:3639 Northern Blot Estrogen stimulates Muc-1 expression. Progesterone inhibits Muc-1 expression. Lane 1 = Day 1 pregnancy Lane 2 = Day 4 pregnancy Day 4 of pregnancy mice ovariectomized and given either P4 (lane 3) or E plus P4 (lane 4). Surveyor et al. 1995, Endocrinology 136:3639 Model for Muc-1 function as an antiadhesion moleucle Surveyor et al. 1995, Endocrinology 136:3639 Role of epithelial surface mucins in steric inhibition of embryo-endometrial interactions Aplin, 1997; Rev. Repod. 2:84-93 Lipid Hydrolysis Prostaglandin Synthesis Increased Membrane Permeability Moulton and Koenig, 1986, In Nidation (ed. Yoshinaga) pp95-109 Heparin Sulfate Proteoglycans av integrin through a bridging ligand Heparan sulfate Cadherin binding to a basic binding protein. Also possibly Chondroitin sulfate Interaction of trophoectoderm and endometrial cells at implantation. Aplin, 1997; Rev. Repod. 2:84-93 Integrins a has 17 subunits b has 8 subunits Endocannabinoid Signaling Endocannabinoids (Marijuana) target receptors (CB1 and CB2) in the mouse embryo. CB1 localized in trophectoderm and not ICM CB1 in oviduct and uterus Knockout of CB1 causes defects in development and survival High levels of the endocannabinoid, Anandamide (AEA), inhibit early embryo development. However, low levels are needed and stimulate trophoblast development. Anandamide levels are also important for implantation. Implantation competency requires down-regulation of AEA binding to the blastocyst. Coordinated down-regulation of CB1 on blastocyst and decrease in AEA from uterus. have Women with elevated peripheral AEA levels spontaneous pregnancy loss. Sun and Dey. Mol Cell Endo (2008) 286:S1-S11 Shelesnyak, 1986, In Nidation (ed. Yoshinaga) pp5-22 Dey and Johnson, 1986, In Nidation (ed. Yoshinaga) pp49-62 Role of Growth Factors in Preimplantation Development IL-6 will bind to LIFr as well Colony Stimulating Factor (CSF-1) has been shown to be involved with implantation too! Leukemia Inhibitor Factor has been demonstrated to be involved with implantation in the mouse. LIF appears following estrogen in delayed implantation LIF stimulates proteinase expression in the uterus. Like - MMPs urokinase type plasminogen activator. LIF not present in delayed implantation Histological features of the uterus in LIFnull and wild-type mice. Semi-thin resin sections from days 5 and 6 of pregnancy were stained with toluidine blue. The insets show high magnification of differentiated polygonal stromal cells in the wild-type and undifferentiated fibroblast-like stroma cells in the LIF-null mice on day 6 of pregnancy. E = embryo, scale bars = 50 µm (Reprinted from Developmental Biology, vol 281, Fouladi-Nashta et al. LIF Activation stimulates Heparin Binding Epidermal Growth Factor Autocrine versus juxtacrine signaling modes. In the simplest model, autocrine signaling is regulated by the removal of the prepro-extension from the membrane-anchored ligand (step 1) followed by its controlled release from the membrane (step 2). Orientation restrictions are responsible for the release requirement. In the case of juxtacrine signaling, prepro-extension release is required (step 1), followed by binding to an auxiliary molecule on a neighboring cell (step 2). Note that autocrine ligands bind to the cell that produced them, and juxtacrine ligands bind to a neighboring cell. Presence of HB-EGF Receptors on Activated Blastocysts Activation of Implantation by LIF induces HB-EGF in LE of Endometrium Amphiregulin HB-EGF Epiregulin Immunofluorescence staining for Cox-2 and oncostatin M (OsM) in LIF-null and wild-type mouse uterus on days 5 and 6 of pregnancy. Cox-2 protein is strongly expressed in the LE and underlying stromal cells (arrows) at the implantation site on day 5 and expression extended deeper into the stroma by day 6. In LIF-null mice, expression was limited to the LE cells and only a few stromal cells expressed Cox-2 in the day-6 uterus. The pattern of expression for OsM protein in wild-type mice was similar to Cox-2. In LIF-null animals, OsM was completely absent around the embryo on days 5 and 6 of pregnancy. E = embryo, scale bar = 100µm (Reprinted from Developmental Biology, vol 281, Fouladi-Nashta et al. Wang and Dey, 2006 Nature Reviews 7:185-199 Kimber (2005) Reproduction 130:131-145 Cross et al. 1994 Science 266:1508 At day 4.5 of mouse development, the blastocyst attaches to the uterine epithelium and uterine lumenal closure occurs. A surge of estrogen from ovary stimulates implantation. Estrogen releases the cytokines such as LIF, IL-1, HB-EGF and CSF-1. Adhesion molecules induce binding through lectincarborhydrate and integrin-integrin interactions Cross et al. 1994 Science 266:1508 Preimplantation Implantation Sherman et al. 1979, In: Matermal Recogniton of Pregnancy, Cibia foundation symposium 64, pp 33-52 Trophoblast cells begin to penetrate epithelium Decidual cells form Implantation Chamber Invasion Phase The uterine epithelium degenerates (apoptosis) and trophoblast cells come into contact with decidua Decidualization - uterine stroma undergoes a transformation, fibroblasts proliferate and enlarge stromal area Postimplantation Phase Trophoblast cells lose their invasiveness Compartive Placentation (ed. D.H. Steven), 1975 Mouse blastocyst implantation Day 7 p.c. Decidual Reaction Note: Position and orientation of blastocyst. Surface epithelium is intact Blastocyst Mouse blastocyst implantation Day 8 p.c. Note: Blastocyst is inside stroma with loss of uterine epithelium Uterine Lumen Perry, 1981; J. Reprod. Fert. 62:321 Tachi and Tachi, 1986, In Nidation (ed. Yoshinaga) pp158-182 Day 5 blastocyst implantation Tung et al. Biol Reprod 35:1045 Tachi and Tachi, 1986, In Nidation (ed. Yoshinaga) pp158-182 Rat Implantation site on Day 7 p.c. Luminal epith Collagen Fibers Decidual cells Tung et al. Biol Reprod 35:1045 Capillary Blastocyst Implantation Chamber Enders and Schlafke, 1979, In: Matermal Recogniton of Pregnancy, Cibia foundation symposium 64, pp 3-32 Tachi and Tachi, 1986, In Nidation (ed. Yoshinaga) pp158-182 Tung et al. Biol Reprod 35:1045 Restriction of dye from the primary decidual zone Tung et al. Biol Reprod 35:1045 Salamonsen, 1999;Rev. Reprod. 4:11-22 Role of Metalloproteinases and Tissue Inhibitor of Metalloproteinases Urokinase-type plasminogen activator Plasminogen MMPs can be regulated by steroids- P4 negative role Cytokines - IL-1, TNF-a MMP secreted in their latent form and most be cleaved to be activated Tissue inhibitor of metalloproteinases a-macroglobulin Salamonsen, 1999;Rev. Reprod. 4:11-22 Development of Fetal Membranes in the Rat Development of the embryo and fetal membranes in the rat is one of space conservation. The conservation of space is achieved through inversion of the germ layers which results in the yolk sac surrounding the embryo. Polar Trophoblast Mural Trophoblast After loss of the zona (D 4-5) the rat blastocyst attaches eccentrically in a groove on the antimesometrial aspect of the uterine wall. The polar trophoblast cells continue to divide to become the ectoplacental cone; these cells engulf and digest maternal erythrocytes. Mural trophoblast cells do not divide, but their DNA content rises as they become transformed into primary giant cells Compartive Placentation (ed. D.H. Steven), 1975 Parietal endoderm Capillary channels in the decidual tissue enter a sinus-like space between the trophoblast and Reichert’s membrane. Because of inversion of germ layers, the Chorion does not surround the embryo. Viseral endoderm The ICM cells continue to increase in number and are pushed down into the blastocoele so that the endoderm surrounds the ectoderm cells (layer inversion) which forms the primary embryonic cavity. A extra-embryonic membrane called Reichert’s membrane appears on Day 6. It first appears as a amorphous secretion along the inner wall of the trophoblast. It lies between the Trophoblastic giant cells and parietal endoderm. Compartive Placentation (ed. D.H. Steven), 1975 Compartive Placentation (ed. D.H. Steven), 1975 Compartive Placentation (ed. D.H. Steven), 1975 Perry, 1981; J. Reprod. Fert. 62:321 Day 7.5 Mouse Embryo Decidual response has occurred. Uterine epithelium have undergone apoptosis. Transformation and growth of stroma cells into decidual cells. Trophoblast giant cells invade the decidua. Invasion regulated by MMP’s and TIMP’s as well as integrins Cross et al. 1994 Science 266:1508 Compartive Placentation (ed. D.H. Steven), 1975 The hemochorial placenta is formed from the ectoplacental cone with its lacunae of maternal blood, in relation to the chorionic ectoderm. Compartive Placentation (ed. D.H. Steven), 1975 Day 9.5 Mouse Embryo Chorioallantoic Placenta Allantois fuses with the chorion. Labyrinthine layer develops in which there is extensive intermingling of maternal blood and fetal blood vessels. Decidua has largely regressed and placental development complete. Cross et al. 1994 Science 266:1508 Stimulus of Implantation 1. The luminal epithelial cells initially differentiate so that their free surface becomes organized is such a way to allow attachment of the trophoblast. 2. During attachment phase, the apposing luminal surfaces of the endometrium come in close contact in all parts of the uterus. Stimulated by estrogen. 3. The blastocyst attachment is antimesometrial with the inner cell mass oriented mesometrially. 4. The blastocyst invades through the uterine epithelium by phagocytosis of the uterine epithelium which has undergone apoptosis. Caused by estrogen not embryo. 5. During invasion process the stroma surrounding the blastocyst has undergone decidualization. Blood vessels in the stroma become more permeable leading to edema with fibroblast cells undergoing waves of cell division with formation of gap junctions between cells. Function of Decidual Cells 1. Barrier against possible immunological rejection of fetus. 2. Protection of the uterus from ravages of the invading trophoblast. 3. Nutritive role until formation of allantois. 4. Endocrine function - estrogen, prolactin, placental lactogen 5. Maternal recognition of pregnancy