Lecture 4 - Susan Schwinning
advertisement

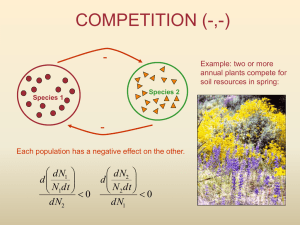
Two-species competition
The Lotka-Volterra Model
Working with differential equations to predict population dynamics
Paramecium caudatum
Testing the
consequences of
species interactions:
Georgii Frantsevich
Gause (b. 1910)
Paramecium aurelia
Gause’s competitive exclusion principle:
Two species competing for the exactly same resources cannot stably
coexist if other ecological factors relevant to the organism remain
constant. One of the two competitors will always outgrow the other,
leading to the extinction of one of the competitors: Complete
competitors cannot coexist.
Overcoming Gause’s exclusion principle:
If two species utilize sufficiently separate niches, the competitive
effects of one species on another decline enough to allow stable
coexistence.
LOTKA AND VOLTERRA
(Pioneers of two-species models)
Alfred J. Lotka (1880-1949)
Chemist, ecologist, mathematician
Ukrainian immigrant to the USA
Vito Volterra (1860-1940)
Mathematical Physicist
Italian, refugee of fascist Italy
LOTKA AND VOLTERRA
(Pioneers of two-species models)
Alfred J. Lotka (1880-1949)
Chemist, ecologist, mathematician
Ukrainian immigrant to the USA
Vito Volterra (1860-1940)
Mathematical Physicist
Italian, refugee of fascist Italy
Let’s say, two species are competing for the same limited space:
In what ways can the species be different?
K1 25
K2 100
The two species might have a different carrying capacities.
time
time
r1 2
per year
r2 4
per year
The two species might have different maximal rates of growth.
When alone each species might follow the logistic
growth model:
For species 1:
K1 N1
dN1
r1 N1
dt
K1
For species 2:
K2 N2
dN2
r2 N 2
dt
K2
When alone each species might follow the logistic
growth model:
For species 1:
K1 N1
dN1
r1 N1
dt
K1
For species 2:
K2 N2
dN2
r2 N 2
dt
K2
How do we express the effect one has on the other?
1 light blue square
has the same effect
as four dark blue
squares.
1 dark blue squares
has the same effect
as 1/4 light blue
square.
The effect of the small purple pecies on the growth rate of the large green species:
N2
1
K1 N1 N 2
dN1
4
r1 N1
dt
K1
N1
The effect of the large orange species on the growth rate of the small blue species:
N1
N2
K 2 N 2 4 N1
dN2
r2 N 2
dt
K2
The Lotka-Volterra two-species competition model:
K1 N1 N 2
dN 1
r1 N1
dt
K1
K 2 N 2 N1
dN2
r2 N 2
dt
K2
Two state variables: N1 and N2, which change in response to one another.
6 parameters: r1, K1, ,r2 ,K2 ,, which stay constant.
and are new to us: they are called interspecific competition coefficients.
The Lotka-Volterra Model is an example of a
system of differential equations:
dN1
f ( N1 , N 2 ,...N m )
dt
dN2
g ( N1 , N 2 ,...N m )
dt
.
.
(differential equations)
dNm
q ( N1 , N 2 ,...N m )
dt
What are the equilibria?
What stability properties do the equilibria have?
Are there complex dynamics and strange attractors for some parameter values?
Analysis tools for systems of two equations:
Isoclines
Definition of the zero-growth isocline:
The set of all {N1,N2} pairs that make the rate of change
for either N1 or N2 equal to zero.
dN1
f ( N1 , N 2 ) 0
dt
dN2
g ( N1 , N 2 ) 0
dt
defines the N1 isocline
defines the N2 isocline
GRAPHICAL ANALYSIS OF TWO-DIMENSIONAL SYSTEMS:
State space graph: a graph with the two state variables on the axes:
Use this graph to plot zerogrowth isoclines, which satisfy:
N2
dN1
0
dt
dN2
0
dt
N1
“N1 isocline”
“N2 isocline”
ISOCLINES:
N2
K1 N1
N2
K1
N2 K2 N1
N1 isocline
K2
N2 isocline
K1
N1
K2
This is called a state space graph.
N2
K1
N1 isocline
K2
The equilibrium!
N2 isocline
K1
N1
K2
The N1 isocline
N2
K1
dN1/(N1dt) < 0
dN1/(N1dt) > 0
N1
K1
The N2 isocline
N2
K2
dN2/(N2dt) < 0
dN2/(N2dt) > 0
N1
K2
This equilibrium is stable!
K1
N2
K2
N1 isocline
K1
N1
N2 isocline
K2
N2
K2
N2 isocline
Case 2:
• an unstable equilibrium
• only one of the two species survives
• which one survives depends on initial
population densities.
K1
N1 isocline
K2
N1
K1
Case 3:
• no two-species equilibrium
• species 1 always wins
K1
N2
K2
N1
K2
K1
Case 4:
• no two-species equilibrium
• species 2 always wins
K2
N2
K1
N1
K1
K2
K1
N2
Case1 :
K2<K1/ and K1<K2/
N2
Case 2:
K2>K1/ and K1>K2/
K1
K2
K1
K1
K2
N1
K2
Case 3:
K2<K1/ and K1>K2/
K2
K2
K1
N1
Case 4:
K2>K1/ and K1<K2/
K1
K2
K2
K1
K1
K2
GENERALIZED STABILITY ANALYSIS
dN1
f ( N1 , N 2 ,...N m )
dt
dN2
g ( N1 , N 2 ,...N m )
dt
.
.
dNm
q ( N1 , N 2 ,...N m )
dt
Step 1: determine all equilibrium points by setting all rates of change to zero
and solve for N.
Step2:
Determine rates of change for each variable at the equilibrium.
Step3:
Determine for every state variable, when in a position just off the
equilibrium, if the are attracted to or repelled by the equilibrium.
Step 1: We rescale equations with respect to the equilibrium of interest:
Define:
x1(t)= N1(t) – N1*
x2(t)= N2(t) – N2* ,
Step 2: We “linearize” the rates of change at the equilibrium:
dx1
a11 x1 a12 x2
dt
dx2
a21 x1 a22 x2
dt
Or, in matrix script:
x Jx
J is called the Jacobian matrix or community matrix in ecology.
Step 3: We find the Jacobian Matrix by finding the partial derivatives of all
differential equations with respect to all state variables:
dN1
f ( N1 , N 2 )
dt
dN2
g ( N1 , N 2 )
dt
Stability identified by determining all partial derivatives, evaluated at the
equilibrium N1*, N2*:
f
a11
N1
a21
g
N1
N1* , N 2*
f
a12
N 2
N1* , N 2*
N1* , N 2*
g
a22
N 2
N1* , N 2*
a11 a12
J
a21 a22
We already know that the eigenvalues of such a matrix can be determined by solving:
lx1 = a11x1+a12x2
lx2 = a21x1+a22x2
As in Leslie matrix analysis, the eigenvalues determine the stability of the equilibrium.
Recall that eigenvalues (roots of polynomials) have the form l = a + bi,
where i = 1
Stability
Real (b=0) Real
and a<0
(b=0) and
a>0
Stable node
l1 and l2
Saddle point
(unstable)
l1
Stable focus
Unstable focus
Linear stability
analysis
insufficient
Complex
(b≠0) and
a<0
Complex
(b≠0) and
a>0
Purely
imaginary
(a=0)
l2
l1 and l2
l1 and l2
l1 and l2
STABLE NODE:
Equilibrium is attracting.
The pathway of approach is monotonic (straight)
N2
N1 isocline
l1 and l2
are both real and
negative
N2 isocline
N1
SADDLE POINT:
Equilibrium is unstable.
The saddle point is attracting in one direction and repelling in another.
N2
N2 isocline
l1 and l2
are both real and one is
negative, the other is
positive
N1 isocline
N1
STABLE FOCUS:
Equilibrium is stable.
The pathway of approach is oscillatory.
N2
N2 isocline
l1 and l2
are complex and the
real part is negative.
N1 isocline
N1
UNSTABLE FOCUS:
Equilibrium is unstable.
The pathway away from the equilibrium is oscillatory.
N2
N2 isocline
l1 and l2
are complex and the
real part is positive.
N1 isocline
N1
NEUTRAL STABILITY:
Equilibrium is neither stable nor unstable.
The pathway is oscillatory and unchanging.
N2
N2 isocline
l1 and l2
are purely imaginary.
N1 isocline
N1
Summary:
1. We search for equilibria to determine the long-term asymptotic behavior of
dynamical systems. This is not limited to population models. We can ask this
about all dynamic models.
2. We use local stability analysis to determine the stability of equilibrium points.
This is done by linearizing the dynamical system near the equilibrium (or near
each equilibrium).
3. The matrix of partial differentials that represent the linearized version of the
dynamical system around a given equilibrium point is called the Jacobian, an
n x n matrix for n differential equations.
4. The eigenvalues of this matrix determine the stability of the equilibrium.