Community Ecology BSC 405
advertisement

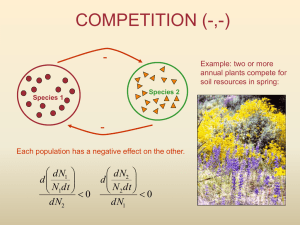
Other patterns in communities • Macroecology: relationships of – geographic distribution and body size – species number and body size • Latitudinal gradients: changes in S with latitude • Species-Area relations: Island biogeography and related questions Species-area relationships • Islands, either oceanic or habitat • Selected areas within continents • How is number of species related to area? S A Mathematics S=c z A – S is number of species – A is area sampled – c is a constant depending on the taxa & units of area – z is a dimensionless constant • often 0.05 to 0.37 Often linearized • ln (S ) and ln (A ) • ln (S ) = ln (c ) + z ln (A ) – z is now the slope – ln (c ) is now the intercept ln (S ) ln (A ) Theory & Hypotheses • Area per se hypothesis – why S goes up with A – why S = c A z – why z takes on certain values • Habitat heterogeneity hypothesis – why S goes up with A • Passive sampling hypothesis – why S goes up with A Area per se Sn • large heterogeneous assemblage log normal distribution of species abundances • assume log normal ("canonical log normal") – Abundance class for most abundant species = abundance class with most individuals – constrains variance (s2) of the distribution • assume that N increases linearly with A • Yield: unique relationship: S = c Az • for "canonical" with S > 20: S = c A0.25 ni Area per se • z varies systematically – larger for real islands vs. pieces of contiguous area • z does not take on any conceivable value – if log normal had s2 = 0.25 (very low) – then z 0.9 … which is virtually unknown in nature – implies constraints on log normal distributions Dynamics of the area per se hypothesis • open island of a given area • rate of immigration (sp. / time) = I initially high • once a species is added, I declines • nonlinear: I – 1st immigrants best dispersers – last are poorest dispersers S ST Dynamics of the area per se hypothesis • rate of extinction (sp. / time) = E initially 0 • as species are added, E increases • nonlinear: E – lower n as S increases – more competition as S increases S ST Dynamic equilibrium RATE • equilibrium when E = I • determines S* • how are rates related to area? E I S S* Effect of area on S* RATE • 2 islands equally far from mainland • large & small • extinction rate greater on small Esmall Elarge I – smaller n’s – greater competition • under this hypothesis I is not related to area S S*small S*large Area per se • Neutral hypotheses vs. Niche hypotheses • Neutral hypotheses – presume that biological and ecological differences between species, though present, are not critical determinants of diversity • Area per se is a neutral hypothesis – S depends only on the equilibrium between species arrival and extinction – Large A large populations low prob. extinction Niche-based hypotheses • Niche hypotheses - presume that that biological and ecological differences between species are the primary determinants of diversity • Niche differences enable species to coexist stably • Does not require equilibrium between extinction and arrival Habitat heterogeneity • Niche-based hypothesis • Larger islands more habitats – Why? • More habitats more species – does not require competition – does not require equilibrium – does not exclude competition or equilibrium • Larger islands bigger “target” • Neutral hypothesis • More immigration more species – competition & equilibrium not necessary (but possible) – under this hypothesis E is not related to area Passive sampling RATE E Ilarge Ismall S S*small S*large Processes • Interspecific competition Competition • Competition occurs when: –a number of organisms use and deplete shared resources that are in short supply –when organisms harm each other directly, regardless of resources –interspecific, intraspecific Resource competition competitor #1 Interference competition competitor #1 + competitor #2 - + - competitor #2 - - resource Competition • Interference – Direct attack – Murder – Toxic chemicals – Excretion • Resource – Food, Nutrients – Light – Space – Water • Depletable, beneficial, & necessary Competition & population • Exponential growth • dN / dt = r N N – r = exponential growth rate – unlimited growth • Nt = N0 ert t Competition & population • Logistic growth: [K-N] dN / dt = r N K • r = intrinsic rate of increase • K = carrying capacity K N t Carrying capacity • Intraspecific competition – among members of the same species • As density goes up, realized growth rate (dN / dt) goes down • What about interspecific competition? – between two different species Lotka-Volterra Competition N1 N2 r1 r2 K1 K2 [ K1 - N1 - a2 N2 ] dN1 / dt = r1 N1 K1 [ K2 - N2 - a1 N1 ] dN2 / dt = r2 N2 K2 Lotka-Volterra Competition • a1 = competition coefficient – Relative effect of species 1 on species 2 • a2 = competition coefficient – Relative effect of species 2 on species 1 • equivalence of N1 and N2 Effects of Ni & Ni’ on growth [ K1 - N1 - a2 N2 ] dN1 / dt = r1 N1 K1 In the numerator, a single individual of N2 has a equivalent effect on dN1 / dt to a2 individuals of N1 Competition coefficients: a’s • Proportional constants relating the effect of one species on the growth of a 2nd species to the effect of the 2nd species on its own growth – a2 > 1 impact of sp. 2 on sp. 1 greater than the impact of sp. 1 on itself – a2 < 1 impact of sp. 2 on sp. 1 less than the impact of sp. 1 on itself – a2 = 1 impact of sp. 2 on sp. 1 equals the impact of sp. 1 on itself Notation dNi / dt vs. • total population growth • dNi / dt = riNi [Ki-Ni-ai’Ni’]/Ki dNi / Nidt • per capita population growth • dNi / Nidt = ri [Ki-Ni-ai’Ni’]/Ki Lotka-Volterra equilibrium • at equilibrium – dN1 / N1dt = 0 & dN2 / N2dt = 0 – also implies dN1 / dt = dN2 / dt = 0, so... • 0 = r1N1 [ (K1-N1-a2N2)/ K1] • 0 = r2N2 [ (K2-N2-a1N1)/ K2] • true if N1 = 0 or N2 = 0 or r1= 0 or r2 = 0 Lotka-Volterra equilibrium • for 0 = r1N1 [ (K1-N1-a2N2)/ K1] • true if 0 = (K1-N1-a2N2)/ K1 • if N2 = 0, implies N1 = K1 (logistic equilibrium) • as N1 0, implies a2N2=K1 or N2 = K1 / a2 • plot as graph of N2 vs. N1 Equilibrium • dNi / dt = 0 for both species • K1 - N1 -a2N2 = 0 and K2 - N2 -a1N1 = 0 N2 Zero Growth Isocline (ZGI) for species 1 K1/a2 dN1/dt<0 dN1/dt>0 N1 K1 Zero growth isocline for sp. 2 N2 dN2 /N2 dt < 0 K2 Zero Growth Isocline (ZGI) dN2 /N2 dt = 0 dN2 /N2 dt > 0 0 K2/a1 N1 Zero growth isocline for sp. 1 N2 dN1 / N1 dt < 0 K1 / a2 Zero Growth Isocline (ZGI) dN1/N1dt = 0 dN1 /N1 dt > 0 0 K1 N1 dN1 / N1dt r1 0 K1 / a2 N2 Isocline in 3 dimensions K1 N1 Zero Growth Isocline ... dN1/N1dt = 0 Isocline in 3 dimensions 0 K1 / a2 N2 K1 N1 Zero Growth Isocline ... dN1/N1dt = 0 N2 K1 / a2 Isocline Zero Growth Isocline ... dN1/N1dt = 0 0 K1 N1 Two Isoclines on same graph • May or may not cross • Indicates whether two competitors can coexist • For equilibrium coexistence, both must have – Ni > 0 – dNi / Ni dt = 0 Lotka-Volterra Zero Growth Isoclines N2 K1/a2 K2 0 K2/a1 Species 1 “wins” K1 N1 • K1 / a2 > K 2 • K1 > K2 / a1 • Region dN1/N1dt>0 & dN2/N2dt>0 • Region dN1/N1dt>0 & dN2/N2dt<0 • Region dN1/N1dt<0 & dN2/N2dt<0 Lotka-Volterra Zero Growth Isoclines N2 K2 K1/a2 0 K1 Species 2 “wins” K2/a1 N1 • K2 > K1 / a2 • K2 / a1 > K1 • Region dN1/N1dt>0 & dN2/N2dt>0 • Region dN1/N1dt<0 & dN2/N2dt>0 • Region dN1/N1dt<0 & dN2/N2dt<0 Competitive Asymmetry • Competitive Exclusion • Suppose K1 K2. What values of a1 and a2 lead to competitive exclusion of sp. 2? • a2 < 1.0 (small) and a1 > 1.0 (large) • effect of sp. 2 on dN1 / N1dt less than effect of sp. 1 on dN1 / N1dt • effect of sp. 1 on dN2 / N2dt greater than effect of sp. 2 on dN2 / N2dt Lotka-Volterra Zero Growth Isoclines N2 K1/a2 • K1 / a2 > K2 • K2 / a1 > K1 • Region both species increase • Regions & one species decreases & one species increases • Region both species decrease Stable coexistence K2 0 K1 K2/a1 N1 Stable Competitive Equilibrium • Competitive Coexistence • Suppose K1 K2. What values of a1 and a2 lead to coexistence? • a1 < 1.0 (small) and a2 < 1.0 (small) • effect of each species on dN/Ndt of the other is less than effect of each species on its own dN/Ndt • Intraspecific competition more intense than interspecific competition Lotka-Volterra Zero Growth Isoclines N 2 K2 • K2 > K1 / a2 Unstable two species equilibrium • K1 > K2 / a1 • Region both species increase • Regions & one species decreases & one species increases • Region both species decrease K1/a2 0 K2/a1 K1 N1 Unstable Competitive Equilibrium • Exactly at equilibrium point, both species survive • Anywhere else, either one or the other “wins” • Stable equilibria at: – (N1 = K1 & N2 = 0) – (N2 = K2 & N1 = 0) • Which equilibrium depends on initial numbers – Relatively more N1 and species 1 “wins” – Relatively more N2 and species 2 “wins” Unstable Competitive Equilibrium • Suppose K1 K2. What values of a1 and lead to coexistence? • a1 > 1.0 (large) and a2 >1.0 (large) • effect of each species on dN/Ndt of the other is greater than effect of each species on its own dN/Ndt • Interspecific competition more intense than intraspecific competition Lotka-Volterra competition • Four circumstances – – – – Species 1 wins Species 2 wins Stable equilibrium coexistence Unstable equilibrium; winner depends on initial N’s • Coexistence only when interspecific competition is weak • Morin, pp. 34-40 Competitive Exclusion Principle • Two competing species cannot coexist unless interspecific competition is weak relative to intraspecific competition • What makes interspecific competition weak? – Use different resources – Use different physical spaces – Use exactly the same resources, in the same place, at the same time Competitve exclusion Model assumptions • All models incorporate assumptions • Validity of assumptions determines validity of the model • Different kinds of assumptions • Consequences of violating different kinds of assumptions are not all the same Simplifying environmental assumption • The environment is, with respect to all properties relevant to the organisms: – uniform or random in space – constant in time • realistic? • if violated need a better experimental system Simplifying biological assumption • All the organisms are, with respect to their impacts on their environment and on each other: – identical throughout the population • clearly must be literally false • if seriously violated need to build a different model with more realistic assumptions Explanatory assumptions • What we propose as an explanation of nature (our hypothesis) – r1, r2, K1, K2, a1, a2 are constants – competition is expressed as a linear decline in per capita growth (dN / N dt ) with increasing N1 or N2 – Some proportional relationship exists between the effects of N1 and N2 on per capita growth • If violated model (our hypothesis) is wrong Interspecific competition: Paramecium • George Gause • P. caudatum goes extinct • Strong competitors, use the same resource (yeast) • Competitve asymmetry • Competitive exclusion Interspecific competition: Paramecium • P. caudatum & P. burseria coexist • P. burseria is photosynthetic • Competitive coexistence • Apparently stable Experiments in the laboratory • • • • • Gause’s work on protozoa Flour beetles (Tribolium) Duck weed (Lemna, Wolffia) Mostly consistent with Lotka Volterra No clear statement of what causes interspecific competition to be weak Alternative Lotka-Volterra competition • Absolute competition coefficients dNi / Nidt = ri [1 – bii Ni - bij Nj] equivalent to: dNi / Nidt = ri [Ki - Ni - aj Nj] / Ki = ri [Ki/Ki - Ni/Ki - ajNj/Ki] = ri [1- (1/Ki)Ni – (aj/Ki)Nj] Absolute Lotka-Volterra N2 1/b12 Stable coexistence 1/b22 0 1/b11 1/b21 N1 Competitive effect vs. response • Effect: impact of density of a species – Self density (e.g., b11) – Other species density (e.g., b21) • Response: how density affects a species – Self density (e.g., b11) – Other species’ density (e.g., b12) • Theory: effects differ (b11 > b21) • Experiments: responses (b11, b12) Absolute Lotka-Volterra N2 1/b12 Stable coexistence 1/b22 0 1/b11 1/b21 N1 Not ecological models • No mechanisms of competition in the model – Phenomenological • Environment not explicitly included • Mechanistic models of Resource competition