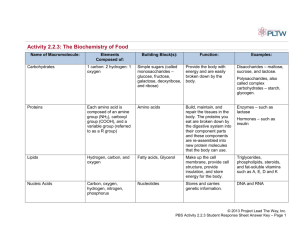
أساسيات الكيمياء الحيوية
advertisement

Principles of BIOCHEMISTRY Dr. AHMED KHAMIS MOHAMED SALAMA Medical Laboratories Dept., Colleges compound at Zulfi February 2013 1 Introduction to Biochemistry The cell is the basic unit of living organisms. The basic components of animal cell are indicated as: 1. Plasma membrane: Surrounds the cell as a selectively permeable membrane. 2. Cytoplasm: Composed mostly of water. 3. Nucleus: Contains genetic information, or instructions for making molecules (DNA). 4. Ribosome: Produces proteins. 5. Lysosome: Contains enzymes that digests wastes. 6. Mitochondrion: Breaks down glucose with oxygen to produce energy. 7. Endoplasmic reticulum: Transports materials throughout the cell. 2 Basic Biochemistry Terms Living systems are composed of very complex molecules (polymers) which built from many small molecules. These small molecules are called monomers and are the basic unit of these polymers. Different kinds and sequences of monomers come together to form compounds with different characteristics. Monomers come together by a process called dehydration synthesis. A hydrogen atom from one functional group joins together with a hydroxide ion from a functional group of another molecule, which results in water and a bond formed between the two molecules. This is called dehydration synthesis. Hydrolysis is just the opposite. 3 Amino acid Amino acids (the monomers of proteins) coming together to make dipeptide. 4 Polarity is very important and plays an important role in cell membranes. You should remember that like dissolves like. Polar molecules will always dissolve in water, while non-polar ones will not. Substances that are attracted to water are called hydrophilic and ones that are not are called hydrophobic. Biomolecules in the living organism: 1. Carbohydrates Carbohydrates are the major energy source for living organisms. Mitochondria break down carbohydrates and use the chemical energy released from its bonds to live. Carbohydrates always have a 1:2:1 ratio of carbon, hydrogen, and oxygen. Since the empirical formula is CH2O, they are called carbohydrates. The monomers of carbohydrates are called monosaccharides and are also called simple sugars. Some popular monosaccharides are glucose, fructose, ribose, and galactose. Oligosaccharides are composed of 2-8 monomers (sucrose, lactose, maltose) . However, some very important carbohydrates are composed of thousands of monomers and are called polysaccharides (starch, cellulose, glycogen). 2. Proteins Proteins provide structure, and organic catalysts are mostly made of proteins. They can also be used to store energy. They are produced by ribosome in our cells. Fibrous proteins provide structure and are usually long and thin. Examples include muscles, hair, cartilage, veins, ducts, and other structures. Globular proteins do things other than structure, like transport oxygen and nutrients, fight invasions by foreign objects, help 5 maintain homeostasis in the body (hormones), and catalyze reactions that would take too long in their absence (enzymes). Conjugated proteins do various functions throughout the body. It means that proteins connected to a non-protein moiety such as a sugar (glycoproteins), nucleic acid (nucleoproteins), lipid (lipoproteins). Amino Acids: They are the monomers of proteins. The basic parts of an amino acid are an amine group, a carboxylic group, and a side chain, all attached to an alpha-carbon (central carbon). The bond between two amino acids is called a peptide bond. There are only 20 R-groups, and therefore only 20 kinds of amino acids found in nature, 8 of which that can be produced by humans and called non-essential amino acids. The rest we have to obtain by eating stuff, and these are called essential amino acids. Surprisingly, the number of combinations of amino acids is almost endless. 6 3. Lipids Lipids are also very important to structure. Lipids and conjugated compounds include fats, oils, phospholipids, waxes, terpens, steroids. A common fat is composed of glycerol and long chains of carboxylic acids called fatty acids. They are generally referred to as triglycerides. Triglycerides can be decomposed by adding NaOH. Fatty acids are saturated if there are no double bonds in their chain, unsaturated if there is one double bond, and polyunsaturated if there is more than one double bond. Saturated fatty acids tend to be solid, since they form straight lines and can stack on top of each other. On the other hand, unsaturated fats are liquid at room temperature because since they have a double bond, their shape is bent and they can't form layers as well. 7 Phospholipids are similar to triglycerides, only the phospholipid has two fatty acid tails and one phosphate group. Our cell membranes are composed of phospholipid bilayers through which small molecules like water and air can diffuse through. Unlike triglycerides and phospholipids, waxes contain monohydroxy alcohols instead of glycerol (which has three hydroxyl groups). They are solids with low melting points, and are used as waterproof both in nature and commercially. Some important steroids 1. Cholesterol: It is the most widespread animal steroid and is found in almost all animal tissues. Cholesterol is a necessary intermediate in the biosynthesis of the steroid hormones; however, since it can be synthesized from acetylcoenzyme-A CH3CO- Co-A , it is not a dietary necessity. High levels of blood cholesterol hardening of the arteries. 2. Cortisone and cortisol These hydrocortisone hormones secreted by the adrenal cortex. They are widely used to treat inflammation due to allergies or rheumatoid arthritis. 3. Sex hormones 8 They are produced primarily in the testes or the ovaries; their production is regulated by pituitary hormones. Male hormones are collectively called androgens; female hormones, estrogens; and pregnancy hormones, progestins. 4. Bile acids They are found in bile, which is produced in the liver and stored in the gall bladder. The structure of cholic acid, the most abundant bile acid, follows. Bile acids are secreted into the intestines to keep lipids emulsified in the intestines, thereby promoting their digestion. 4. Nucleic Acids In order for life to go on for generations, organisms need to transmit genetic material to the next generation. Somehow this information needs to be stored somewhere, and be easy to duplicate during reproduction. DNA and RNA have the perfect structure for governing the cell (telling it what molecules to make and how) and are practical to duplicate. 9 Nucleotides: are the monomers of nucleic acids. They are made up of three parts: a five-carbon sugar, a nitrogenous base, and a phosphate group: 10 11 1. CARBOHYDRATES Carbohydrates are the most abundant class of biomolecules found in living organisms. They originate as products of photosynthesis, an endothermic reductive condensation of carbon dioxide requiring light energy and the pigment chlorophyll. 12 Carbohydrates fill numerous roles in living things, such as: Carbohydrates are the major source of metabolic energy (storage and transport of energy, starch, glycogen). Carbohydrates serve as a structural components (cellulose in plants, chitin in animals). Carbohydrates are one of the three essential components of DNA and RNA. Carbohydrates and their derivatives play major roles in the working process of the immune system, pathogenesis, blood clotting, and development. The formulas of many carbohydrates can be written as carbon hydrates, Cn(H2O)n. Many carbohydrates have the empirical formula CH2O; for example, the molecular formula for glucose is C6H12O6 (six times CH2O). So carbohydrates are naturally occurring compounds of carbon, hydrogen, and oxygen. 13 Carbohydrates are actually polyhydroxy aldehydes and ketones or their derivatives. The carbohydrates vary dramatically in their properties. For example, sucrose (table sugar) and cellulose (plant) are both carbohydrates. One of the principal differences between various types of carbohydrates is the size of the molecules. The monosaccharides (often called simple sugars) are the simplest carbohydrate units; they cannot be hydrolyzed to smaller carbohydrate molecules. Monosaccharides can bonded together to form dimmer, trimer, etc. and, ultimately, polymers. The dimmers are called disaccharides. Sucrose is a disaccharide that can be hydrolyzed to one unit of glucose plus one unit of fructose. The monosaccharides and disaccharides are soluble in water and are generally sweet-tasting. 14 15 The names of carbohydrates often end in the suffix -ose. Carbohydrates composed of 2 – 8 units of monosaccharides are referred to as oligosaccharides. If more than 8 units of monosaccharides result from hydrolysis, the carbohydrate is a polysaccharide. Example of polysaccharides are: starch, found in flour and cornstarch. Cellulose, a fibrous constituent of plants and the principal component of cotton. 16 The following figure shows dimentional formulas and Fischer projections for monosaccharides. 17 Fischer projections: Emil Fischer introduced projection formulas for showing the spatial arrangement of groups around chiral carbon atoms. Dimensional formula of the chain The Fischer projection of the chain form of D-glucose form of D-glucose 18 Fischer projection is simply a shorthand way to represent a ball – and – stick or dimensional formula. In drawing a Fischer projection, we assume that the molecule is completely stretched out in the plane of the paper with all its substituent. Each intersection of the horizontal and vertical lines represents a chiral carbon atom. Each horizontal line represents a bond coming toward the viewer, while the vertical line represents bonds going back, away from the viewer. A pair of enantiomers of pentose is easily recognized when Fischer projections are used. Mirror image 19 Fischer projections Emil Fischer introduced projection formulas for showing the spatial arrangement of groups around chiral carbon atoms. Nomenclature Aldoses such as glucose (six carbon atoms) are named aldohexose, ripose (five carbon atoms) are named aldopentose and so on. Ketoses such as fructose (six carbon atoms) are named hexulose, xylose (five carbon atoms) are named pentulose and so on. 20 Aldose Aldotriose D-Glyceraldehyde Aldotetroses D-Erythrose D-Threose 21 Aldopentoses D-Ribose D-Arabinose D-Xylose D-Lyxose Aldohexoses D-Allose D-Altrose D-Glucose D-Mannose 22 D-Gulose D-Idose D-Galactose D-Talose Ketoses Triulose Dihydroxyacetone Tetrulose D-Erythrulose 23 Pentulose D-Ribulose D-Xylulose Hexulose D-Psicose D-Fructose 24 D-Sorbose D-Tagatose SOME MONOSACCHARIDES Glucose: Glucose is the most important of the monosaccharides, is sometimes called blood sugar (because it is found in the blood), grape sugar (because it is found in grapes), or dextrose (because it is dextrorotatory). Mammals can convert sucrose, lactose (milk sugar), maltose, and starch to glucose, which is then used by the organism for energy or stored as glycogen (a polysaccharide). When the organism needs energy, the glycogen is again converted to glucose. Excess carbohydrates can be converted to fat; therefore a person can become obese on a fat-free diet. Carbohydrates can also be converted to steroids (such as cholesterol) and, to a limited extent, to protein. Conversely, an organism can convert proteins and fats to carbohydrates. 25 Fructose: Fructose is ketosugars, have sex carbon atoms, named hexulose. It is also called levulose because it is levorotatory. Fructose is the sweetest-tasting of all the sugars. It occurs in fruit and honey, as well as in sucrose. 26 Galactose Galactose is aldosugars, have sex carbon atoms, aldohexoses. It is found in milk as it is named bonded to glucose, in the disaccharide lactose. Ribose and deoxyribose Ribose is form part of the polymeric backbone of the nucleic acid RNA while deoxyribose is form part of the polymeric backbone of the nucleic acid DNA. The prefix deoxy- means “ minus an oxygen”; the structures of ribose and deoxyribose are the same except that deoxyribose lacks an oxygen at carbon 2. 27 Classification of monosaccharides Monosaccharides are classified according to: 1. The placement of its carbonyl group If the carbonyl group is an aldehyde, the monosaccharide is an aldose; if the carbonyl group is a ketone, the monosaccharide is a ketose. Examples for aldoses: glucose, galactose, ribose, and deoxyribose. Examples for ketoses: fructose 28 2. The number of carbon atoms it contains (usually from 3-7 atoms) Monosaccharides with three carbon atoms are called trioses, those with four are called tetroses, five are called pentoses, six are hexoses, and so on. These two systems of classification are often combined. For example, glucose is an aldohexose (a six-carbon aldehyde), ribose is an aldopentose (a five-carbon aldehyde), and fructose is a ketohexose (a six-carbon ketone). 3. Its optical isomers. Each carbon atom bearing a hydroxyl group (-OH) are asymmetric, making them stereo-centers. The assignment of D or L is made according to the orientation of the asymmetric carbon furthest from the carbonyl group: in a standard Fischer projection if the hydroxyl group is on the right the molecule is a D sugar, otherwise it is an L sugar. Because D sugars are biologically far more common, the D is often omitted. 29 Configuration of Glucose Glucose has six carbon atoms, four of which are chiral centers (Carbons 2,3,4, and 5) indicate there may be as many as sixteen (24) stereoisomers. Only half of these sixteen sterioisomers belong to the D-series; of these, only D-glucose, D-galactose, and D-mannose occur in abundance. The Fisher projection of all the D-aldoses, from D-glyceraldehyde through the D-aldohexoses, are shown below. The addition of one more carbon creates a new chiral carbon in each step down in the figure. Therefore, D-glyceraldehyde leads to a pair of 30 tetroses, each tetrose leads to a pair of pentoses and each pentose leads to a pair of hexoses. 31 Ketoses 32 33 Cyclization of Monosaccharides The preferred structural form of many monosaccharides may be that of a cyclic hemiacetal. A general reaction of alcohols and aldehydes is that of hemiacetal formation Five and six-membered rings are favored over other ring sizes because of their low angle and eclipsing strain. Cyclic structures of this kind are termed furanose (five-membered) or pyranose (six-membered), reflecting the ring size relationship to the common heterocyclic compounds furan and pyran. 34 During the conversion from straight-chain form to cyclic form, the carbon atom containing the carbonyl oxygen, called the anomeric carbon, becomes a chiral center with two possible configurations: the oxygen atom may take a position either above or below the plane of the ring. The resulting possible pair of stereoisomers are called anomers (- anomer and β-anomer). In the - anomer, the -OH substituent on the anomeric carbon rests on the opposite side (trans) of the ring from the CH2OH side branch. The alternative form, in which the CH2OH substituent and the anomeric hydroxyl are on the same side (cis) of the plane of the ring, is called the β-anomer. Because the ring and straight-chain forms readily interconvert, both anomers exist in equilibrium. In water solution, glucose can undergo an intramolecular reaction to yield cyclic hemiacetals. Either five-membered ring hemiacetals (using 35 the hydroxyl group at carbon 4) or six-membered ring hemiacetals (using the hydroxyl group at carbon 5) can be formed. 36 Ribose, an important aldopentose, commonly adopts a furanose structure, as shown in the following illustration. By convention for the D-family, the five-membered furanose ring is drawn in an edgewise projection with the ring oxygen positioned away from the viewer. The anomeric carbon atom (colored red here) is placed on the right. The upper bond to this carbon is defined as beta, the lower bond then is alpha. Note that carbon 1 (the aldehyde carbon), which is not chiral in the open-chain structure, because chiral in the cyclization. Therefore, a pair of diastereomers results from the cyclization (called anomers). A monosaccharide in the form of a five-membered ring hemiacetal is called a furanose. Similarly, the six-membered ring form is called a pyranose after pyran. The terms furanose and pyranose are often combined with the name of the mono-saccharide- forexample, Dglucopyranose for the six-membered ring of D-glucose, or Dfructofuranose for the five-membered ring of fructose. The cyclic pyranose forms of various monosaccharides are often drawn in a flat projection known as a Haworth formula. 37 The terminal- CH2OH group is positioned above the plane of the ring in the D-series and below the plane of the ring in the L-series. 38 Note that any group that is to the right in the Fischer projection is down in the Haworth projection, and any group that is to the left in the Fischer projection is up in the Haworth formula. Mutarotation: Pure glucose exists in two crystalline forms: -D-glucose and -Dglucose. Pure -D-glucose has a melting point of 146 C. The specific rotation of a freshly prepared solution is +112. Pure -D-glucose has a melting point of 150 C. and a specific rotation of +18.7. The specific rotation of a solution of either - or -D-glucose changes 39 slowly until it reaches an equilibrium value of +52.6. This slow spontaneous change in optical rotation was called mutarotation. 40 Glycosides When a hemiacetal is treated with an alcohol, an acetal is formed. The acetals of monosaccharides are called glycosides and have names ending in -oside. 41 Important reactions of Carbohydrates Oxidation of monosaccharides: As noted above, sugars may be classified as reducing or non-reducing based on their reactivity with Tollens', Benedict's or Fehling's reagents. If a sugar is oxidized by these reagents it is called reducing, since the oxidant (Ag+ or Cu+2) is reduced in the reaction, as evidenced by formation of a silver mirror or precipitation of cuprous oxide. The Tollens' test is commonly used to detect aldehyde functions; and because of the facile inter-conversion of ketoses and aldoses under the basic conditions of this test, ketoses such as fructose also react and are classified as reducing sugars. When the aldehyde function of an aldose is oxidized to a carboxylic acid the product is called an aldonic acid. If both ends of an aldose chain are oxidized to carboxylic acids the product is called an aldaric acid. Thus, ribose, xylose, allose and galactose yield achiral aldaric acids which are, of course, not optically active. The ribose oxidation is shown in equation 2 below. 1. Aldonic acid 42 2. Aldaric acid 3. Uronic acids: 43 4. Periodic acid oxidation: The periodic acid oxidation is a test for 1,2-diols and for 1,2- or hydroxy aldehydes and ketones. A compound containing such a grouping is oxidized and cleaved by periodic acid. The products of the periodic acid oxidation of glucose are formaldehyde and formic acid in the molar ration of 1:5 This oxidation of the CHOH group in glucose to formic acid can be rationalized by considering the reaction to be stepwise. 44 5. Effect of concentrated acids on monosaccharides: Concentrated acids caused dehydration for the monosaccharide leading to furfural in case of pentoses or 5-hydroxy methyl furfural in case of hexoses. 45 6. Reduction of monosaccharides: 7. Chain Shortening and Lengthening These two procedures permit an aldose of a given size to be related to homologous smaller and larger aldoses. 46 7. Esterification: 8. Acetate formation: A common reagent for esterification of alcohols is acetic anhydride, with either sodium acetate or pyridine as an alkaline catalyst. 47 9. Ether formation: Dimethyl sulfate is an inorganic ester with an excellent leaving group. This compound is used to form methyl ethers. Disaccharides A disaccharides is a carbohydrate composed of two units of monosaccharide joined together by a covalent bond known as a glycoside link from carbon 1 of one unit to an OH of the other unit (via a dehydration reaction, resulting in the loss of a hydrogen atom from one monosaccharide and a hydroxyl group from the other). A common mode of attachment is an or glycoside link from the first unit to the 4-hydroxy group of the second unit. This link is called a 1,4-- or a 1,4- link, depending on the stereochemistry at the glycoside carbon. The formula of unmodified disaccharides is C12H22O11. Although there are numerous kinds of disaccharides, a handful of disaccharides are particularly notable. 48 Sucrose: Sucrose is the most abundant disaccharide and the main form in which carbohydrates are transported in plants. The disaccharide sucrose is common table sugar. Whether it comes from beets or sugar cane, the chemical composition of sucrose is the same. It is composed of two monosaccharides: D-glucose and D- fructose. The glycoside link joins the ketal and acetal carbons; from fructose and from glucose. The systematic name for sucrose, O-α-D-glucopyranosyl-(1→2)-Dfructofuranoside, indicates four things: 1. Its monosaccharides: glucose and fructose. 2. Their ring types: glucose is a pyranose, and fructose is a furanose. 3. How they are linked together: the oxygen on carbon number 1 (C1) of -D-glucose is linked to the C2 of D-fructose. 4. The -oside suffix indicates that the anomeric carbon of both monosaccharides participates in the glycosidic bond. Note the difference between sucrose and the other disaccharide is that both anomeric carbon atoms (not just one) are used in the glycoside link. In sucrose, neither fructose nor glucose has a hemiacetal group; 49 therefore, sucrose in water is not in equilibrium with an aldehyde or keto form. Sucrose does not exhibit mutarotation and is not a reducing sugar. Invert sugar is a mixture of D-glucose and D-fructose obtained by the acidic or enzymatic hydrolysis of sucrose. The enzymes that catalyze the hydrolysis of sucrose, called invertases. Honey is primarily invert sugar. Maltose: The disaccharides maltose is used in baby foods and malted milk. It is the principal disaccharide obtained from the hydrolysis of starch. Starch is broken down into maltose in an apparently random fashion by an enzyme in saliva called -1,4-glucan 4-glucanohydrolase. A molecule of maltose contains two units of D-glucopyranose. The first unit is in the form of an -glycoside. This unit is attached to the oxygen at carbon 4- in the second unit by a 1,4-- link. 50 Cellobiose: The disaccharide obtained from the partial hydrolysis of cellulose is called cellobiose. Like maltose, cellobiose is composed of two glucopyranose units joined together by a 1,4 - link. Cellobiose differs from maltose in that the 1,4-linkage is β- rather than -. 51 Lactose: The disaccharide lactose (milk sugar) is different from maltose or cellobiose in that it is composed of two different monosaccharides, Dglucose and D-galactose. The systematic name for lactose is O-β-D-galactopyranosyl-(1→4)-Dglucopyranose. 52 Lactose is naturally occurring disaccharide found only in mammal molecule, occurs naturally in milk. The systematic name for lactose is O-β-D-galactopyranosyl-(1→4)-D-glucopyranose.s; human milk contain about 5% lactose. cow’s Lactose milk is and obtained commercially as a by-product in the manufacture of cheese. In normal human metabolism, lactose is hydrolyzed enzymatically to D-galactose and D-glucose; then the galactose is converted to glucose, which can undergo metabolism. A condition called GALACTOSEMIA that affects some infants is caused by lack of the enzyme used to concert galactose to glucose. Galactosemia is characterized by high levels of galactose in the blood and urine. Symptoms ranges from vomiting to mental and physical retardation and sometimes death. Treatment consists of removing milk and milk products from the diet. (An artificial milk made from soybeans may be substituted. Polysaccharides: A polysaccharid is a compound in which the molecules contain many units of monosaccharid joined together by glycoside links. Upon complete hydrolysis, a polysaccharide yields monosaccharides. Polysaccharides serve three purposes in living systems: 1. Architectural purpose: cellulose gives strength to the plant stems and branches, chitin serve as structural component of the exoskeletons of insects. 2. Nutritional purpose: starch is found in wheat and potatoes, glycogen is an animal’s internal store of readily available carbohydrate. 3. Specific agents: heparin prevents blood coagulation. 53 Oligosaccharides and polysaccharides are composed of longer chains of monosaccharide units bound together by glycosidic bonds. The distinction between the two is based upon the number of monosaccharide units present in the chain. Oligosaccharides typically contain between two and nine monosaccharide units, and polysaccharides contain greater than ten monosaccharide units. Definitions of how large a carbohydrate must be to fall into each category vary according to personal opinion. Starch: Starch is a polymer of glucose, found in roots, rhizomes, seeds, stems, tubers and corms of plants, as microscopic granules having characteristic shapes and sizes. Most animals, including humans, depend on these plant starches for nourishment. Starch consists of two fractions. About 20% is a water soluble material called amylose. Molecules of amylose are linear chains of several thousand glucose units joined by alpha C-1 to C-4 glycoside bonds. Amylose solutions are actually dispersions of hydrated helical micelles. The majority of the starch is a much higher molecular weight substance, consisting of nearly a million glucose units, and called amylopectin (80%). Molecules of amylopectin are branched networks built from C-1 to C-4 and C-1 to C-6 glycoside links, and are essentially water insoluble. Amylose is a linear polymer of glucose mainly linked with α-1,4 bonds. It can be made of several thousands of glucose units. It is one of the two components of starch, the other being amylopectin. 54 Hydrolysis of starch, usually by enzymatic reactions, produces a syrupy liquid consisting largely of glucose. When cornstarch is the feedstock, this product is known as corn syrup. It is widely used to soften texture, add volume, prohibit crystallization and enhance the flavor of foods. Glycogen: Glycogen is the glucose storage polymer used by animals. It has a structure similar to amylopectin, but is even more highly branched (about every tenth glucose unit). The degree of branching in these polysaccharides may be measured by enzymatic or chemical analysis. 55 Cellulose and chitin: Cellulose and chitin are examples of structural polysaccharides. Cellulose is used in the cell walls of plants and other organisms, and is claimed to be the most abundant organic molecule on earth. It has a variety of uses including in the paper and textile industry and as a feedstock for the production of rayon (in the viscose process), cellulose acetate, celluloid and nitrocellulose. Chitin has a similar structure to cellulose but has nitrogen containing side branches, increasing its strength. It is found in arthropod exoskeletons and in the cell walls of some fungi. It has a variety of uses, for example in surgical threads. 56 Over half of the total organic carbon in the earth's biosphere is in cellulose. Cotton fibres are essentially pure cellulose, and the wood of bushes and trees is about 50% cellulose. As a polymer of glucose, cellulose has the formula (C6H10O5)n where n ranges from 500 to 5,000, depending on the source of the polymer. The glucose units in cellulose are linked in a linear fashion. Most animals cannot digest cellulose as a food, and in the diets of humans this part of our vegetable intake functions as roughage and is eliminated largely unchanged. Some animals (the cow and termites, for example) harbor intestinal microorganisms that breakdown cellulose into monosaccharide nutrients by the use of beta-glycosidase enzymes. 57 Cellulose is commonly accompanied by a lower molecular weight, branched, amorphous polymer called hemicellulose. In contrast to cellulose, hemicellulose is structurally weak and is easily hydrolyzed by dilute acid or base. Also, many enzymes catalyze its hydrolysis. Hemicelluloses are composed of many D-pentose sugars, with xylose being the major component. Mannose and mannuronic acid are often present, as well as galactose and galacturonic acid. Unlike cellulose, hemicellulose (also a polysaccharide) consists of shorter chains - 500-3,000 sugar units as opposed to 7,000 - 15,000 glucose molecules per polymer seen in cellulose. In addition, hemicellulose is a branched polymer, while cellulose is unbranched. Study problems 1. Draw a Haworth formula of anomers of D-glucopyranose and D-fructofuranose ? 58 2. There are eight D- aldohexoses and eight L-aldohexoses. Write Fischer and Haworth progections of D- and L- glucose. Haworth projection for D- and L-glucose 59 Fischer projection for D- and L- glucose 3. Oxidation of monosaccharides gives different compounds, e.g. aldonic acid, aldaric acid or uronic acid. Explain ? 60 4. Classify the following monosaccharides (as an aldohexose, e.g.) and give the number of isomers of each: 61 5. Write equation that illustrate the MUTAROTATION of pure D-glucose in water ? 62 6. Give the structure of the products for the followings: D-Glucose + HIO4 D-Glucose + [H] β-D-Glucose + (CH3 CO)2O NaO2CCH3 H2O Starch H2O …… ……. ……………. H+ or enzymes H+ or enzymes 7. Draw 1,4- Ethanol enzymes α-glycoside link in maltose ? 7. What is the difference between each pair of the following? Lactose and sucrose Maltose and cellobiose Amylose and amylopectin 63 2. LIPIDS Lipids are broadly defined as any fat-soluble naturally-occurring molecules, such as fats, oils, waxes, cholesterol, sterols, vitamins A, D, E and K, glycerols, phospholipids, and others. 64 Biological functions of lipids: 1. Acting as structural components of cell membranes. Phospholipids are lipids that contain phosphate ester groups. The phospholipids are the main structural component of biological membranes, such as the cellular plasma membrane and the intracellular membranes of organelles. In animal cells the plasma membrane physically separates the intracellular components from the extracellular environment. Phospholipids are made from glycerol, two fatty acids, and (in place of the third fatty acid) a phosphate group with some other molecule attached to its other end (e.g. acetyl choline or ethanol amine). The hydrocarbon tails of the fatty acids are hydrophobic, but the phosphate group end of the molecule is hydrophilic. This means that phospholipids are soluble in both water and oil. A biological membrane is a form of lipid bilayers. In an aqueous system, the polar heads of lipids orientate towards the polar, aqueous environment, while the hydrophobic tails minimize their contact with water. Cell membranes are made mostly of phospholipids arranged in 65 a double layer with the tails from both layers “inside” (facing toward each other) and the heads facing environment) on both surfaces. 66 “out” (toward the watery 2. Energy storage and metabolism. Triacylglycerols, stored in adipose tissue, are a major form of energy storage in animals. Animals use triglycerides for energy storage because of its high caloric content (9 KCal/g). Triglycerides and phospholipids are broken down into free fatty acids by the action of lipases. Beta oxidation is the process by which fatty acids, in the form of acyl-CoA molecules, are broken down in the mitochondria and/or in peroxisomes to generate acetyl-CoA. The acetyl CoA is then ultimately converted into ATP, CO2 , and H2O using the citric acid cycle and the electron transport chain. Conversely, fatty acid biosynthesis takes place in the cytoplasm, using acetyl-CoA (derived from carbohydrates, amino acids or fatty acids) as the precursor. The fatty acids may be subsequently converted to triacylglycerols that are packaged in lipoproteins and secreted from the liver. Many lipids are absolutely essential for life. However, there is also considerable awareness: Abnormal levels of certain lipids, particularly cholesterol (in hypercholesterolemia) and trans fatty acids, are risk factors for heart disease amongst others. Humans have a requirement for certain essential fatty acids, such as linoleic acid (omega-6 fatty acid) and α-linolenic acid (omega-3 fatty acid) in the diet because they cannot be synthesized from simple precursors in the diet. Both of these fatty acids are 18-carbon polyunsaturated fatty acids differing in the number and position of the double bonds. Most vegetable oils are rich in linoleic acid (sunflower, and corn oils). α-linolenic acid is found in the green leaves of plants, and in 67 selected seeds, nuts and legumes (flax, canola, walnuts and soy). Fish oils are particularly rich in the longer-chain omega-6 fatty acids eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). Most of the lipid found in food is in the form of triacylglycerols, cholesterol and phospholipids. Most of the saturated fatty acids (as triacylglycerols) in the diet are incorporated into adipose tissue stores, because the absence of double bonds allows a higher energy yield per carbon than is obtained from oxidation of unsaturated fatty acids. The longer chain fatty acids are incorporated into cell membranes as phospholipids regardless of degree of saturation. Dietary fat provides an average energy intake which is approximately twice that of carbohydrate or protein. A minimum amount of dietary fat is necessary to facilitate absorption of fat-soluble vitamins (A, D, E and K) and carotenoids. A minimal amount of body fat is also necessary to provide insulation that prevents heat loss and protects vital organs from shock due to ordinary activities. High fat intake contributes to increased risk of obesity, diabetes and atherosclerosis. Atherosclerosis is the primary cause of coronary and cardiovascular diseases and is primarly due to the buildup of plaque on the inside walls of arteries. Plaque is made up of cholesterol-rich low density lipoproteins (LDL). Saturated fats have a profound hypercholesterolemic (increase blood cholesterol levels) effect and tend to increase plasma LDL. They are found predominantly in animal products (butter, cheese and meat). 68 Intake of monounsaturated fats in oils such as olive oil is thought to be preferable to consumption of polyunsaturated fats in oils such as corn oil because the monounsaturated fats apparently do not lower high-density-lipoprotein (HDL) cholesterol levels. Keeping cholesterol in the normal range not only helps prevent heart attacks and strokes but may also prevent the progression of atherosclerosis. Lipids are consists of the following: 1. Fats and oils 2. waxe 3. phospholipids 4. steroids (like cholesterol) 5. some other related compounds. Fats and Oils Fats and oils are made from two kinds of molecules: glycerol (a type of alcohol with a hydroxyl group on each of its three carbons) and three fatty acids joined by dehydration synthesis. Since there are three fatty acids attached, these are known as triglycerides. Triglycerides or triacylglycerol 69 The distinction between a fat and oil is arbitrary (random): at room temperature a fat is solid and an oil is liquid. Most glycerides in animals are fats, while those in plants tend to be oil; hence the terms animal fats (beef fat) and vegetable oils (corn oil, sunflower oil). The carboxylic acid obtained from the hydrolysis of a fat or oil, called a fatty acid, generally has a long, unbranched hydrocarbon chain. Fats and oils are often named as derivatives of these fatty acids. Most naturally occurring fats and oils are mixed triglycerides- that is, the three fatty-acid portions of the glyceride are not the same. Almost all naturally occurring fatty acids have an even number of carbon atoms because they are biosynthesized from the two-carbon acetyl groups in acetylcoenzyme A. 70 The rate of fatty acid synthesis is controlled by the equilibrium between monomeric acetyl Co-A carboxylase (ACC) and polymeric ACC. The activity of ACC requires polymerization. This conformational change is enhanced by citrate and inhibited by long-chain fatty acids. The synthesis of fatty acids from acetyl-CoA and malonyl-CoA is carried out by fatty acid synthase (FAS). The primary fatty acid synthesized by fatty acid synthase (FAS) is palmitate enzyme CH3 (CH2)14 COO- . and can then Palmitate is then released from the undergo separate unsaturation to yield other fatty acid molecules. Structure of Fatty Acids 71 elongation and/or The “tail” of a fatty acid is a long hydrocarbon chain, making it hydrophobic. The “head” of the molecule is a carboxyl group which is hydrophilic. Fatty acids are the main component of soap, where their tails are soluble in oily dirt and their heads are soluble in water to emulsify and wash away the oily dirt. However, when the head end is attached to glycerol to form a fat, that whole molecule is hydrophobic. The terms saturated, mono-unsaturated, and poly-unsaturated refer to the number of hydrogens attached to the hydrocarbon tails of the fatty acids as compared to the number of double bonds between carbon atoms in the tail. Fats, which are mostly from animal sources, have all single bonds between the carbons in their fatty acid tails, thus all the carbons are also bonded to the maximum number of hydrogen possible. Since the fatty acids in these triglycerides contain the maximum possible amount of hydrogen, these would be called saturated fats. The hydrocarbon chains in these fatty acids are, thus, fairly straight and can pack closely together, making these fats solid at room temperature. 72 Oils, mostly from plant sources, have some double bonds between some of the carbons in the hydrocarbon tail, causing bends or “kinks” in the shape of the molecules. Because some of the carbons share double bonds, they’re not bonded to as many hydrogen’s as they could if they weren’t double bonded to each other. Therefore these oils are called unsaturated fats. Because of the kinks in the hydrocarbon tails, unsaturated fats can’t pack as closely together, making them liquid at room temperature. Many people have heard that the unsaturated fats are “healthier” than the saturated ones. Hydrogenated vegetable oil (as in shortening and commercial peanut butters where a solid consistency is needed) started out as “good” unsaturated oil. However, this commercial product has had all the double bonds artificially broken and hydrogen’s artificially added to turn it into saturated fat that bears no resemblance to the original oil from which it came (so it will be solid at room temperature). In unsaturated fatty acids, there are two ways the pieces of the hydrocarbon tail can be arranged around a C=C double bond. In cis bonds, the two pieces of the carbon chain on either side of the double bond are either both “up” or both “down,” such that both are on the same side of the molecule. 73 In trans bonds, the two pieces of the molecule are on opposite sides of the double bond, that is, one “up” and one “down” across from each other. Naturally-occurring unsaturated vegetable oils have almost all cis bonds, but using oil for frying causes some of the cis bonds to convert to trans bonds. If oil is used only once like when you fry an egg, only a few of the bonds do this so it’s not too bad. However, if oil is constantly re-used, like in fast food French fry machines, more and more of the cis bonds are changed to trans until significant numbers of fatty acids with trans bonds build up. The reason this is of concern is that fatty acids with trans bonds are carcinogenic, or cancer-causing. The levels of trans fatty acids in highly-processed, lipid-containing products such as margarine are quite high. We need fats in our bodies and in our diet. Animals in general use fat for energy storage because fat stores 9 KCal/g of energy. Plants, which don’t move around, can afford to store food for energy in a less compact but more easily accessible form, so they use starch for energy storage. 74 Carbohydrates and proteins store only 4 KCal/g of energy, so fat stores over twice as much energy/gram as fat. By the way, this is also related to the idea behind some of the high-carbohydrate weight loss diets. The human body burns carbohydrates and fats for fuel in a given proportion to each other. The theory behind these diets is that if they supply carbohydrates but not fats, then it is hoped that the fat needed to balance with the sugar will be taken from the dieter’s body stores. Fat is also is used in our bodies to: Cushion (soften) vital organs like the kidneys. Serve as insulation, especially just beneath the skin. Examples Of Fatty Acids: 75 The higher melting points of the saturated fatty acids (solid) reflect the uniform rod-like shape of their molecules. The cis-double bond(s) in the unsaturated fatty acids introduce a kink (bend) in their shape, which makes it more difficut to pack their molecules together in a stable repeating array or crystalline lattice. The trans-double bond isomer of oleic acid, known as elaidic acid, has a linear shape and a melting point of 45 ºC (32 ºC higher than its cis isomer). The shapes of stearic and oleic acids are shown below. 76 The configuration around any double bond in a naturally occurring fatty acid is cis, a configuration that results in the low melting points of oils. A saturated fatty acid forms zigzag chains that can fit compactly together, resulting in high Van der Waals attractions; therefore, saturated fats are solids. If a few cis double bonds are present in the chains, the molecules cannot form neat, compact lattices, but tend to be oil. ; polyunsaturated triglycerides tend to be oils. Soaps and Detergents Soaps are the alkali metal salts (usually sodium salts) of fatty acids. Carboxylic acids and salts having alkyl chains longer than eight carbons exhibit unusual behavior in water due to the presence of both hydrophilic (CO2) and hydrophobic (alkyl) regions in the same molecule. Such molecules are termed amphiphilic (Gk. amphi = both) or amphipathic. Fatty acids made up of ten or more carbon atoms are nearly insoluble in water, and because of their lower density, float on the surface when mixed with water. Unlike paraffin or other alkanes, which tend to (pond or spot)) puddle on the waters surface, these fatty acids spread evenly over an extended water surface, eventially forming a monomolecular layer in which the polar carboxyl groups are hydrogen bonded at the water interface, and the hydrocarbon chains 77 are aligned together away from the water. Substances that accumulate at water surfaces and change the surface properties are called surfactants. 78 Alkali metal salts of fatty acids are more soluble in water than the acids themselves, and the amphiphilic character of these substances also make them strong surfactants. The most common examples of such compounds are soaps and detergents, four of which are shown below. Note that each of these molecules has a nonpolar hydrocarbon chain, the "tail", and a polar (often ionic) "head group". The use of such compounds as cleaning agents is facilitated by their surfactant character, which lowers the surface tension of water, allowing it to penetrate and wet a variety of materials. Very small amounts of these surfactants dissolve in water to give a random dispersion of solute molecules. 79 However, when the concentration is increased an interesting change occurs. The surfactant molecules reversibly assemble into poly-molecular aggregates called micelles. By gathering the hydrophobic chains together in the center of the micelle, disruption of the hydrogen bonded structure of liquid water is minimized, and the polar head groups extend into the surrounding water where they participate in hydrogen bonding. These micelles are often spherical in shape, but may also assume cylindrical and branched forms. Here the polar head group is designated by a blue circle, and the nonpolar tail is a zig-zag black line. An animated display of micelle formation is presented below. Micelles are able to encapsulate non-polar substances such as grease within their hydrophobic center, and thus solubilize it so it is removed with the wash water. Since the micelles of anionic amphiphiles have a negatively charged surface, they repel one another and the non-polar dirt is effectively emulsified. To summarize, the presence of a soap or a detergent in water facilitates the wetting of all parts of the object to be cleaned, and removes water-insoluble dirt by incorporation in micelles. 80 The oldest amphiphilic cleaning agent known to humans is soap. Soap is manufactured by the base-catalyzed hydrolysis (saponification) of animal fat. Fat is heated with lye (sodium hydroxide) and is thus saponified to glycerol and sodium salts of fatty acids. Before sodium hydroxide was commercially available, a boiling solution of potassium carbonate leached from wood ashes was used. Soft potassium soaps were then converted to the harder sodium soaps by washing with salt solution. The importance of soap to human civilization is documented by history, but some problems associated with its use have been recognized. One of these is caused by the weak acidity (pKa ca. 4.9) of the fatty acids. Solutions of alkali metal soaps are slightly alkaline (pH 8 to 9) due to hydrolysis. If the pH of a soap solution is lowered by acidic contaminants, insoluble fatty acids precipitate and form a scum. A second problem is caused by the presence of calcium and magnesium 81 salts in the water supply (hard water). These divalent cations cause aggregation of the micelles, which then deposit as a dirty scum. These problems have been alleviated by the development of synthetic amphiphiles called detergents (or syndets). By using a much stronger acid for the polar head group, water solutions of the amphiphile are less sensitive to pH changes. Also the sulfonate functions used for virtually all anionic detergents confer greater solubility on micelles incorporating the alkaline earth cations found in hard water. Variations on the amphiphile theme have led to the development of other classes, such as the cationic and nonionic detergents shown above. Cationic detergents often exhibit germicidal properties, and their ability to change surface pH has made them useful as fabric softners and hair conditioners. These versatile chemical "tools" have dramatically transformed the household and personal care cleaning product markets over the past fifty years. 82 Phospholipids Phospholipids are lipids that contain phosphate ester groups. They are the main constituents of cell membranes. 83 Phospholipids are made from glycerol, two fatty acids, and (in place of the third fatty acid) a phosphate group with some other molecule attached to its other end. The hydrocarbon tails of the fatty acids are still hydrophobic, but the phosphate group end of the molecule is hydrophilic because of the oxygen’s with all of their pairs of unshared electrons. This means that phospholipids are soluble in both water and oil. Phosphoglycerides , one type of phospholipids, are closely related to the fats and oils. These compounds usually contain fatty-acid esters at two positions of glycerol with a phosphate ester at the third position. Phosphoglycerides are distinctive (unique) because their molecules contain two long hydrophobic tails and a highly polar hydrophilic group – a diploar-ion group. Phosphoglycerides are, therefore, neutral surfactants. They are excellent emulsifying agents. In mayonnaise, the 84 phosphoglycerides of the egg yolk keep the vegetable oil emulsified in the vinegar. There are two types of phosphoglyceride (lecithins and cephalins) that are found principally in the brain, nerve cells, and liver of animals and are also found in egg yolks, wheat germ, yeast, soybeans, and other foods. These two types of compounds are similar to each other in structure. 1. LECITHINS (Phosphatidylcholine) are derivatives of choline chloride, HOCH2CH2N(CH3)3 + Cl- , which is involved in the transmission of nerve impulses. 2. CEPHALINS (Phosphatidylethanolamine) are derivatives of ethanolamine, HOCH2CH2NH2. Other classes of phospholipids are represented by plasmalogens, which have vinyl ether group instead of ester groups at carbon 1 of glycerol, and sphingolipids, of which sphingomyelin is an example. 85 Phospholipids aggregate or self-assemble when mixed with water, but in a different manner than the soaps and detergents. Because of the two pendant alkyl chains present in phospholipids and the unusual mixed charges in their head groups, micelle formation is unfavorable relative to a bilayer structure. If a phospholipid is smeared over a small hole in a thin piece of plastic immersed in water, a stable planar bilayer of phospholipid molecules is created at the hole. As shown in the following diagram, the polar head groups on the faces of the bilayer contact water, and the hydrophobic alkyl chains form a nonpolar interior. The phospholipid molecules can move about in their half the bilayer, but there is a significant energy barrier preventing migration to the other side of the bilayer. 86 This bilayer membrane structure is also found in aggregate structures called liposomes. Liposomes are microscopic vesicles consisting of an aqueous core enclosed in one or more phospholipid layers. They are formed when phospholipids are vigorously mixed with water. Unlike micelles, liposomes have both aqueous interiors and exteriors. Terpenes Compounds classified as terpenes constitute what is arguably the largest and most diverse class of natural products. A majority of these compounds are found only in plants, but some of the larger and more complex terpenes ( e.g. squalene & lanosterol ) occur in animals. Terpenes incorporating most of the common functional groups are known, so this does not provide a useful means of classification. Instead, the number and structural organization of carbons is a definitive characteristic. Terpenes may be considered to be made up of isoprene ( more accurately isopentane ) units, an empirical feature known as the 87 isoprene rule. Because of this, terpenes usually have 5n carbon atoms ( n is an integer ). Terpenes are subdivided as follows: Isoprene itself, a C5H8 gaseous hydrocarbon, is emitted by the leaves of various plants as a natural byproduct of plant metabolism. Next to methane it is the most common volatile organic compound found in the armosphere. Examples of C10 and higher terpenes, representing the four most common classes are shown in the following diagram. The initial display is of monoterpenes; larger terpenes will be shown. Most terpenes may be structurally dissected into isopentane segments. 88 The isopentane units in most of these terpenes are easy to discern, and are defined by the shaded areas. In the case of the monoterpene camphor, the units overlap to such a degree it is easier to distinguish them by coloring the carbon chains. This is also done for alpha-pinene. In the case of the triterpene lanosterol we see an interesting deviation from the isoprene rule. This thirty carbon compound is clearly a terpene, and four of the six isopentane units can be identified. However, the ten carbons in center of the molecule cannot be disected in this manner. Polymeric isoprenoid hydrocarbons have also been identified. Rubber is undoubtedly the best known and most widely used compound of this kind. It occurs as a colloidal suspension called latex in a number of plants, ranging from the dandelion to the rubber tree (Hevea brasiliensis). Rubber is a polyene, and exhibits all the expected reactions of the C=C function. Bromine, hydrogen chloride and hydrogen all add with a stoichiometry of one molar equivalent per 89 isoprene unit. Ozonolysis of rubber generates a mixture of levulinic acid ( CH3COCH2CH2CO2H ) and the corresponding aldehyde. Pyrolysis of rubber produces the diene isoprene along with other products. The double bonds in rubber all have a Z-configuration, which causes this macromolecule to adopt a kinked or coiled conformation. This is reflected in the physical properties of rubber. Despite its high molecular weight (about one million), crude latex rubber is a soft, sticky, elastic substance. Chemical modification of this material is normal for commercial applications. Gutta-percha (structure above) is a naturally occuring E-isomer of rubber. Here the hydrocarbon chains adopt a uniform zig-zag or rod like conformation, which produces a more rigid and tough substance. Uses of gutta-percha include electrical insulation and the covering of golf balls. 90 Pheromones Humans communicate by talking, using sign language, painting pictures, or writing letters (or books). But insects and some animals also communicate, in part, by chemicals. 91 A chemical secreted by one individual of a species the brings forth a response in another individual of the same species is called a pheromone. Insect pheromones called alarm pheromones signify danger. For example, a pheromone secreted by one bee helps alert other bees to the location of a food source. Insect sex attractants are another class of pheromones. Sex-attractant pheromones have been used for insect control. In some cases, male insects may be lured by a sex-attractant pheromone, trapped, and then sterilized and released to mate unproductively with the females. Many insect pheromones are not complex in structure. Geranitol and citral, both terpenes, are recruiting pheromones for honeybees, while isoamyl acetate (not a terpene) is a bee alarm pheromone. Steroids The important class of lipids called steroids are actually metabolic derivatives of terpenes, but they are customarily treated as a separate group. 92 Steroids may be recognized by their tetracyclic skeleton, consisting of three fused six-membered and one five-membered ring. The four rings are designated A, B, C & D as noted, and the peculiar numbering of the ring carbon atoms (shown in red) is the result of an earlier misassignment of the structure. The substituents designated by R are often alkyl groups, but may also have functionality. The R group at the A:B ring fusion is most commonly methyl or hydrogen, that at the C:D fusion is usually methyl. The substituent at C-17 varies considerably, and is usually larger than methyl if it is not a functional group. The most common locations of functional groups are C-3, C-4, C-7, C-11, C-12 & C-17. Ring A is sometimes aromatic. Since a number of tetracyclic triterpenes also have this tetracyclic structure, it cannot be considered a unique identifier. Some important steroids 1. Cholesterol It is the most widespread animal steroid and is found in almost all animal tissues. Human gallstones and egg yolks are especially rich sources of this compound. Cholesterol is a necessary intermediate in the biosynthesis of the steroid hormones; however, since it can be synthesized from acetylcoenzyme A, it is not a dietary necessity. High levels of blood cholesterol are associated with arteriosclerosis (hardening of the arteries), a condition in which cholesterol and other lipids coat the insides of the arteries. 2. Cortisone and cortisol These hydrocortisone are two of 28 or more hormones secreted by the adrenal cortex. Both these steroids alter protein, carboxylate, and lipid 93 metabolism in ways not entirely understood. They are widely used to treat inflammation due to allergies or rheumatoid arthritis. 3. Sex hormones They are produced primarily in the testes or the ovaries; their production is regulated by pituitary hormones. The sex hormones impart secondary sex characteristics and regulate the sexual and reproductive functions. Male hormones are collectively called androgens; female hormones, estrogens; and pregnancy hormones, progestins. 94 4. Bile acids They are found in bile, which is produced in the liver and stored in the gall bladder. The structure of cholic acid, the most abundant bile acid, follows.Bile acids are secreted into the intestines in combination with sodium salts of either glycine or taurine. The bile acid-amino acid link is an amide link between the carboxyl group of the bile and the amino group of the amino acid. In this combined form, the bile acid-amino acid acts to keep lipids emulsified in the intestines, thereby promoting their digestion. 95 Lipid Soluble Vitamins The essential dietary substances called vitamins are commonly classified as "water soluble" or "fat soluble". Water soluble vitamins, such as vitamin C, are rapidly eliminated from the body and their dietary levels need to be relatively high. The lipid soluble vitamins are not as easily eliminated and may accumulate to toxic levels if consumed in large quantity. They are vitamin A, vitamin D, vitamin E and vitamin K. From this data it is clear that vitamins A and D, while essential to good health in proper amounts, can be very toxic. Vitamin D, for example, is used as a rat poison, and in equal weight is more than 100 times as poisonous as sodium cyanide. 96 From the structures shown here, it should be clear that these compounds have more than a solubility connection with lipids. Vitamins A is a terpene, and vitamins E and K have long terpene chains attached to an aromatic moiety. The structure of vitamin D can be described as a steroid in which ring B is cut open and the remaining three rings remain unchanged. The precursors of vitamins A and D have been identified as the tetraterpene beta-carotene and the steroid ergosterol, respectively. Waxes Waxes are esters of fatty acids with long chain monohydric alcohols (one hydroxyl group). Natural waxes are often mixtures of such esters, and may also contain hydrocarbons. The formulas for three well known waxes are given below, with the carboxylic acid moiety and the alcohol moiety. 97 Waxes are widely distributed in nature. The leaves and fruits of many plants have waxy coatings, which may protect them from dehydration and small predators. The feathers of birds and the fur of some animals have similar coatings which serve as a water repellent. Carnuba wax is valued for its toughness and water resistance. 98 Study problems: 1. Compare between each pair of the followings: Fat Oil Pheromones Sex hormones Sesquiterpenes Monoterpenes 2. Draw the structure of the followings: The ring system of all steroids Dipeptide Isoprene unit Neutral amino acid One of the terpenes One of the pheromones 99 Phosphoglyceride Uronic acid 3-Match each of the following on the right: 1 Cholesterol Is a steroid hormone 2 Testosterone Phosphoglycerides 3 Pheromones Lipid 4 Corn oil The monomer of terpenes 5 Lecithin Fatty acid 6 Isoprene Synthesized from acetyl Co-A 7 Oleic acid Geranitol 100 3. PROTEINS Proteins are large organic compounds made of amino acids arranged in a linear chain and joined together by peptide bonds between the carboxyl and amino groups of adjacent amino acid residues. Proteins are polyamides, and hydrolysis of a protein yields amino acids. 101 Like other biological macromolecules such as polysaccharides and nucleic acids, proteins are essential parts of organisms and participate in every process within cells. Many proteins are enzymes that catalyze biochemical reactions and are vital to metabolism. Proteins also have structural or mechanical functions, such as actin and myosin in muscle and the proteins in the cytoskeleton, which form a system of scaffolding that maintains cell shape. Other proteins are important in cell signaling, immune responses, cell adhesion, and the cell cycle. Proteins are also necessary in animals' diets, since animals cannot synthesize all the amino acids they need and must obtain essential amino acids from food. Through the process of digestion, animals break 102 down ingested protein into free amino acids that are then used in metabolism. Proteins are linear polymers built from 20 different -amino acids. These amino acids are commonly found in plant and animal proteins, yet these 20 amino acids can be combined in a variety of ways to form muscles, tendons, skin, fingernails, feathers, silk, hemoglobin, enzymes, antibodies, and many hormones. The structure of amino acids: The amino acids found in proteins are -amino carboxylic acids. All amino acids possess common structural features, including an -carbon to which an amino group, a carboxyl group, and a variable side chain are bonded. Only proline differs from this basic structure as it contains an unusual ring to the N-end amine group, which forces the CO–NH amide moiety into a fixed conformation. The side chains of the standard amino acids, detailed in the list of standard amino acids, have different chemical properties that produce three-dimensional protein structure and are therefore critical to protein function. The amino acids in a polypeptide chain are linked by peptide bonds formed in a dehydration reaction. Once linked in the protein chain, an individual amino acid is called a residue, and the linked series of carbon, nitrogen, and oxygen atoms are known as the main chain or protein backbone. 103 Due to the chemical structure of the individual amino acids, the protein chain has directionality. The end of the protein with a free carboxyl group is known as the C-terminus or carboxy terminus, whereas the end with a free amino group is known as the N-terminus or amino terminus. The commonly occurring amino acids are of 20 different kinds which contain the same dipolar ion group H3N+.CH.COO-. They all have in common a central carbon atom to which are attached a hydrogen atom, an amino group (NH2) and a carboxyl group (COOH). The central carbon atom is called the Calpha-atom and is a chiral centre. All amino acids found in proteins encoded by the genome have the Lconfiguration at this chiral centre. Amino acids in proteins (or polypeptides) are joined together by peptide bonds. The sequence of R-groups along the chain is called the primary structure. 104 The simplest amino acid is amino acetic acid (H2NCH2CO2H), called glycine, which has no side chain and consequently does not contain a chiral carbon. All other amino acids have side chains, and therefore their -carbons are chiral. Isomerism Of the standard -amino acids, all but glycine can exist in either of two optical isomers, called L- or D-amino acids, which are mirror images of each other. While L-amino acids represent all of the amino acids found in proteins during translation in the ribosome, D-amino acids are found in some proteins produced by enzyme post-translational modifications after translation and translocation to the endoplasmic reticulum, as in exotic sea-dwelling organisms such as cone snails. They are also abundant components of the peptidoglycan cell walls of bacteria, and D-serine may act as a neurotransmitter in the brain. The L and D convention for amino acid configuration refers not to the optical activity of the amino acid itself, but rather to the optical activity of the isomer of glyceraldehyde from which that amino acid can 105 theoretically be synthesized (D-glyceraldehyde L-glyceraldehyde is levorotary). The two optical isomers 106 is dextrorotary; Amino acids from proteins belong the L-series- that is, the groups around the -carbon have the same configuration as L-glyceraldehyde. Amino acids do not always behave like organic compounds because: 1. They have melting points of over 200 , whereas most organic compounds of similar molecular weight are liquids at room temperature. 2. Amino acids are soluble in water and other polar solvents, but insoluble in non-polar solvents such as diethyl ether or benzene. 3. Amino acids have large dipole moments. 4. They are less acidic than most carboxylic acids and less basic than most amines. Why do amino acids exhibit such unusual properties? The reason is that an amino acid contains a basic amino group and an acidic carboxyl group in the same molecule. An amino acid undergoes an internal acidbase reaction to yield a dipolar ion, also called a zwitter ion. 107 Because of the resultant ionic charges, an amino acid has many properties of a salt. Furthermore, the pKa of an amino acid is not the pKa of a –COOH group, but that of an -NH3 + group. The pKb is not that of a basic amino group, but that of the very weakly basic –COOgroup. 108 ISOELECTRIC POINT Amino acids have both amine and carboxylic acid functional groups and are therefore both an acid and a base at the same time. At a certain pH known as the isoelectric point an amino acid has no overall charge, since the number of protonated ammonia groups (positive charges) and deprotonated carboxylate groups (negative charges) are equal. 109 The amino acids all have different isoelectric points. The ions produced at the isoelectric point have both positive and negative charges and are known as a zwitter ion, which comes from the German word Zwitter meaning "hermaphrodite" or "hybrid". Amino acids can exist as zwitter ions in solids and in polar solutions such as water, but not in the gas phase. Zwitter ions have minimal solubility at their isolectric point and an amino acid can be isolated by precipitating it from water by adjusting the pH. Essential and non-essential amino acids Amino acids are join together to form short polymer chains called peptides or longer chains called either polypeptides or proteins. These polymers are linear and un-branched, with each amino acid within the chain attached to two neighboring amino acids. When taken up into the human body from the diet, amino acids are either used to synthesize proteins and other biomolecules or oxidized to urea and carbon dioxide as a source of energy. The oxidation pathway starts with the removal of the amino group by a transaminase, the amino group is then fed into the urea cycle. The other product of transamidation is -keto acid that enters the citric acid cycle. Of the twenty amino acids, eight are called essential amino acids because the human body cannot synthesize them from other compounds at the level needed for normal growth, so they must be obtained from food. However, the rest can be synthesized in our body and therefore named non-essential amino acids. 110 Individual differences in requirements for amino acids: The amino acid tyrosine is conditionally non-essential where in most people, tyrosine can be synthesized from phenylalanine. A small percent of individuals who have inherited a condition known as phenylketonuria (PKU), which means “phenyl ketones in the urine,” do not have the enzyme phenylalanine hydroxylase necessary for this conversion. In these people, the phenylalanine is converted to phenylpyruvic acid. The diet of a person with PKU must contain some tyrosine and must also be limited in its quantity of phenylalanine; otherwise, excessive amounts of phenylpyruvic acid accumulate in the brain, causing mental retardation. 111 The structure of the common amino acids found in proteins are: 112 Importance of side chain: There are three types of amino acids: 1. Acidic amino acids: They contain more carboxyl groups in their side chain. 2. Basic amino acids: They contain more amino groups in their side chain. 113 3. Neutral amino group: They contain groups other than acidic or amino in their side chains. They divided into two categories: a)-Polar neutral amino acids: They contain –OH, -SH, or other polar groups b)-Non-polar neutral amino acids: They contain non-polar groups. 114 This side chains of amino acids determine the structure reactivity and the characteristic of the protein. The order in which amino acids are found in a protein molecule determines the relationship of the side chains to one another and consequently determines how the protein interacts with itself and with its environment. For example , a hormone or other water-soluble protein contains many amino acids with polar side chains, while an insoluble muscle protein contains a greater proportion of amino acids with nonpolar side chains. Synthesis of amino acids: 1) Amination of alpha-bromocarboxylic acids: The bromoacids, in turn, are conveniently prepared from carboxylic acids by reaction with Br2 + PCl3. Although this direct approach gave mediocre results when used to prepare simple amines from alkyl halides, it is more effective for making amino acids, thanks to the reduced nucleophilicity of the nitrogen atom in the product. Nevertheless, more complex procedures that give good yields of pure compounds are often chosen for amino acid synthesis. 2) By modifying the nitrogen as a phthalimide salt: This procedure, known as the Gabriel synthesis, can be used to advantage in aminating bromo malonic esters. Since the phthalimide substituted malonic ester has an acidic hydrogen, activated by the two ester groups, this intermediate may be converted to an ambident anion and alkylated. Finally, base catalyzed hydrolysis of the phthalimide moiety and the esters, followed by acidification and thermal decarboxylation, produces an amino acid and phthalic acid (not shown). 115 3.An elegant procedure, known as the Strecker synthesis: Assembles an alpha-amino acid from ammonia, cyanide and an aldehyde. This reaction (shown below) is essentially an imino analog of cyanohydrin formation. The alpha-amino nitrile formed in this way can then be hydrolyzed to an amino acid by either acid or base catalysis. 3. Resolution The racemic amino acid is first converted to a benzamide derivative to remove the basic character of the amino group. Next, an ammonium salt is formed by combining the carboxylic acid with an optically pure amine, such as brucine (a relative of strychnine). 116 Reaction of amino acids: 1. Amphoterism of amino acids: As amino acids have both a primary amine group and a primary carboxyl group, therefore, an amino acid is amphoteric: it can undergo reaction with either an acid or a base to yield a cation or an anion, respectively. 2. Acylation of amino acids: The amino group of an amino acid can be readily acylated with either an acid halide or an acid anhydride to yield an amide. 117 3. Reaction with ninhydrin: Amino acids react with ninhydrin to form a blue-violet product called Ruhemann’s purple. The reaction is commonly used as a spot test to detect the presence of amino acids on chromatography paper. 118 4.Carboxylic acid esterification: 5.Oxidative coupling: Peptide bond formation The condensation of two amino acids to form a peptide bond are carried as described below: 119 As both the amine and carboxylic acid groups of amino acids can react to form amide bonds, one amino acid molecule can react with another and become joined through an amide linkage. This polymerization of amino acids is what creates proteins. This condensation reaction yields the newly formed peptide bond and a molecule of water. Chemical synthesis of proteins Short proteins can also be synthesized chemically by a family of methods known as peptide synthesis, which rely on organic synthesis techniques such as chemical legation to produce peptides in high yield. Chemical synthesis allows for the introduction of non-natural amino acids into polypeptide chains, such as attachment of fluorescent probes to amino acid side chains. These methods are useful in laboratory biochemistry and cell biology, though generally not for commercial applications. Chemical synthesis is inefficient for polypeptides longer than about 300 amino acids, and the synthesized proteins may not readily assume their native tertiary structure. Most chemical synthesis methods proceed from C-terminus to N-terminus, opposite the biological reaction. Structure of proteins Most proteins fold into unique 3-dimensional structures. The shape into which a protein naturally folds is known as its native state. Biochemists often refer to four distinct aspects of a protein's structure (Higher structure of proteins): 1. Primary structure: The sequence of amino acids in a protein molecule is called the primary structure of the protein. 120 The primary structure of a segment of a polypeptide chain or of a protein is the amino-acid sequence of the polypeptide chain(s), without regard to spatial arrangement (apart from configuration at the alphacarbon atom). 2. Secondary structure: regularly repeating local structures stabilized by hydrogen bonds. The most common examples are the alpha helix and beta sheet. Because secondary structures are local, many regions of different secondary structure can be present in the same protein molecule. The common secondary structures in proteins are namely -helix, -sheets and turns. Anti-parallel beta-sheets are more often twisted than parallel sheets. Turns are the third of the three "classical" secondary structures that serve to reverse the direction of the polypeptide chain. They are located primarily on the protein surface and accordingly contain polar and charged residues. 121 3. Tertiary structure: the overall shape of a single protein molecule; the spatial relationship of the secondary structures to one another. Tertiary structure is generally stabilized by non-local interactions, most commonly the formation of a hydrophobic core, but also through salt bridges, hydrogen bonds, disulfide bonds, and even 122 post-translational modifications. The term "tertiary structure" is often used as synonymous with the term fold. The tertiary structure of a protein molecule, or of a subunit of a protein molecule, is the arrangement of all its atoms in space, without regard to its relationship with neighboring molecules or subunits. The folding of the backbone upon itself to form a sphere, are called the tertiary structure. 4. Quaternary structure: the shape or structure that results from the interaction of more than one protein molecule, usually called protein subunits in this context, which function as part of the larger assembly or protein complex. The quaternary structure of a protein molecule is the arrangement of its subunits in space and the ensemble of its intersubunit contacts and interactions, without regard to the internal geometry of the subunits. The subunits in a quaternary structure must be in non-covalent association. Haemoglobin contains four polypeptide chains held 123 together non-covalently in a specific conformation as required for its function. Classes of Proteins: Proteins can be informally divided into three main classes, which correlate with typical tertiary structures: 1.Globular proteins: These are small proteins, somewhat spherical in shape because of folding of the protein chains upon themselves. Globular proteins are water-soluble and perform various functions in an organism. For example, hemoglobin transports oxygen to the cells; insulin aids in carbohydrates metabolism; antibodies render foreign protein inactive; fibrinogen (soluble) can form insoluble fibers that result in blood clots; and hormones carry messages throughout the body. Examples of globular proteins are albumines, globulines and histones. 124 2.Fibrous proteins (structural proteins): They are composed of long thread-like molecules that are tough and insoluble. It forms skin, muscles, the walls of arteries and hair. Examples of fibrous proteins are collagens, elastins and keratins. 3.Conjugated proteins (membrane proteins): Proteins connected to a non-protein moiety such as a sugar, perform various functions throughout the body. A common mode of action linkage between the protein and non-protein is by a functional side chain of the protein. For example, an acidic side chain of the protein can form an ester with an –OH group of a sugar molecule. Examples of conjugated proteins are nucleoproteins, mucoproteins, glycoproteins and lipoproteins. Almost all globular proteins are soluble and many are enzymes. Fibrous proteins are often structural; conjugated proteins often serve as receptors or provide channels for polar or charged molecules to pass through the cell membrane. A special case of intramolecular hydrogen bonds within proteins, poorly shielded from water attack and hence promoting their own dehydration, are called dehydrons. Cellular functions The best-known role of proteins in the cell is their duty as enzymes, which catalyze chemical reactions. Enzymes are usually highly specific 125 catalysts that accelerate only one or a few chemical reactions. Enzymes carry out most of the reactions involved in metabolism and catabolism, as well as DNA replication, DNA repair, and RNA synthesis. Some enzymes act on other proteins to add or remove chemical groups in a process known as post-translational modification. About 4,000 reactions are known to be catalyzed by enzymes. The rate acceleration conferred by enzymatic catalysis is often enormous - as much as 1017fold increase in rate over the un-catalyzed reaction in the case of orotate decarboxylase (78 million years without the enzyme, 18 milliseconds with the enzyme). The molecules bound and acted upon by enzymes are known as substrates. Although enzymes can consist of hundreds of amino acids, it is usually only a small fraction of the residues that come in contact with the substrate, and an even smaller fraction - 3-4 residues on average - that are directly involved in catalysis. The region of the enzyme that binds the substrate and contains the catalytic residues is known as the active site. Protein Denaturation Protein denaturation has been defined in several ways, for example as a change in solubility or by simultaneous changes in chemical, physical and biological properties under some standard reference set of conditions. These changes in physical, and to a lesser extent chemical properties are manifestations of configurational changes taking place in the polypeptide chains. The denaturation process presumably involves an unfolding or at least an alteration in the nature of the folded structure. Most denaturation 126 changes consist of changes in secondary bonds: ion-dipole, hydrogen and Van der Waals, and in the rotational positions about single bonds which are controlled by the secondary bond structure. The term denaturation denotes the response of the native protein to heat, acid, alkali, and a variety of other chemical and physical agents which cause marked changes in the protein structure. Rice et al. (1958) suggested denaturation to mean a class of reactions which lead to changes in the structure of the macromolecule with no change in molecular weight. Timasheff and Gibbs (1957), pointed out that the approaches used to define the concept of denaturation can be classified into two types: molecular, in terms of actual structural changes taking place on the molecule, and operational, in terms of changes in measurable properties. The operational approach to denaturation has the advantage of being purely phenomenological, but it cannot lead to a precise definition, 127 since the concept of property, even restricted to measurable property, is itself without finite boundary. On the other hand, the molecular point of view, although still involves a few assumptions despite the important advances in investigations of protein structure, will allow us to reach a definition which is very convenient, owing to its general validity. Whatever it is, changes can be quantitatively described by comparison with the native state. The denaturation process can be achieved by any one of the following methods: Increasing temperature. Changing pH. Using denaturants (i.e. urea, guanidine hydrochloride, betamercaptoethanol, dithiothreitol). Using inorganic salts (i.e lithium bromide, potassium thiocyanate, sodium iodide). Using organic solvents and (i.e. formamide, dimethylformamide, dichloro- and trichloroacetic acids and their salts). Using detergents (i.e. sodium dodecyl sulphate). High pressure. Ultrasonic homogenization. 128 Enzymes Enzymes are biological catalysts responsible for supporting almost all of the chemical reactions that maintain animal homeostasis. Because of their role in maintaining life processes, the assay and pharmacological regulation of enzymes have become key elements in clinical diagnosis and therapeutics. The macromolecular components of almost all enzymes are composed of protein, except for a class of RNA modifying catalysts known as ribozymes. Ribozymes are molecules of ribonucleic acid that catalyze reactions on the phosphodiester bond of other RNAs. Enzymes are found in all tissues and fluids of the body. Almost every significant life process is dependent on enzyme activity. Intracellular enzymes catalyze the reactions of metabolic pathways. Plasma membrane enzymes regulate catalysis within cells in response to extracellular signals. Circulatory system enzymes are responsible for regulating the clotting of blood. Enzyme Classifications Traditionally, enzymes were simply assigned names by the investigator who discovered the enzyme. As knowledge expanded, systems of enzyme classification became more comprehensive and complex. Currently enzymes are grouped into six functional classes by the International Union of Biochemists (I.U.B.). 129 The enzyme's name is comprised of the names of: The substrate(s), The product(s) and The enzyme's functional class. In the enzyme acetyl choline esterase for example, It catalyzes the breakdown of the neurotransmitter acetylcholine at several types of synapses as well as at the neuromuscular junction — the specialized synapse that triggers the contraction of skeletal muscle. One molecule of acetylcholinesterase breaks down 25,000 molecules of acetylcholine each second. This speed makes possible the rapid "resetting" of the synapse for transmission of another nerve impulse. The substrate of this enzyme is acetyl choline. The products are acetate and choline base. 130 The enzyme functional clase is esterase because it is hydrolyze the ester bond in the acetyl choline. Classification of Enzymes Classification of enzymes- More than 2000 different enzymes are currently known. The commonly used names for most enzymes describe the type of reaction catalyzed, followed by the suffix -ase. For example, dehydrogenases hydrolyze proteins, and remove isomerases hydrogen atoms, proteases catalyze rearrangements in configuration. Modifiers may precede the name to indicate the substrate (xanthine oxidase), the source of the enzyme (pancreatic ribonuclease), its regulation (hormone-sensitive lipase) etc. To address ambiguities, the International Union of Biochemists (IUB) developed an unambiguous system of enzyme nomenclature in which each enzyme has a unique name and code number that identify the type of reaction catalyzed and the substrates involved. Enzymes are grouped into six classes: 1) The oxidoreductases (class 1) catalyze the transfer of reducing equivalents(Hydrogen and electrons)from one redox system to another. 2) The transferases (class 2) catalyze the transfer of other groups from one molecule to another. Oxidoreductases and transferases generally require coenzymes 3) The hydrolases (class 3) hydrolases cause cleavage of bond using water 4) Lyases (class 4, often also referred to as“synthases”) catalyze reactions involving either the cleavage or formation of chemical bonds, 131 with double bonds either arising or disappearing.(See figure- reversible reaction is shown). Cleavage of bond does not require water. 5) The isomerases (class 5) move groups within a molecule, without changing the gross composition of the substrate. 6) The ligation reactions catalyzed by ligases (“synthetases,” class 6) are energy-dependent and are therefore always coupled to the hydrolysis of nucleoside triphosphates(See figure) Each enzyme is entered in the Enzyme Catalogue with a four-digit Enzyme Commission number (EC number). The first digit indicates membership of one of the six major classes. The next two indicate subclasses and subsubclasses. The last digit indicates where the enzyme belongs in the subsubclass. For example, The IUB name of hexokinase is ATP:D-hexose 6phosphotransferase E.C. 2.7.1.1. This name identifies hexokinase as a member of class 2 (transferases), subclass 7 (transfer of a phosphoryl group), sub-subclass 1 (alcohol is the phosphoryl acceptor), and “hexose-6″ indicates that the alcohol phosphorylated is on carbon six of a hexose. However, it is still called as hexokinase. 132 Figure- Showing the classification of enzymes with examples of each class of enzymes: Enzymes are also classified on the basis of their composition: 1. Simple enzymes: They are composed wholly of protein. 133 2. Complex enzymes: They are composed of protein plus non-protein component (a relatively small organic molecule). Complex enzymes are also known as HOLOENZYMES. In this terminology the protein component is known as the APOENZYME. Non-protein component is known as the COENZYME or PROSTHETIC GROUP. When the binding between the apoenzyme and non-protein components is non-covalent, the small organic molecule is called coenzyme. When the binding between the apoenzyme and non-protein components is covalent, the small organic molecule is called Prosthetic group. Many prosthetic groups and coenzymes are water-soluble derivatives of vitamins. It should be noted that the main clinical symptoms of dietary vitamin insufficiency generally arise from the malfunction of enzymes , which lack sufficient cofactors derived from vitamins to maintain homeostasis. The non-protein component of an enzyme may be as simple as a metal ion or as complex as a small non-protein organic molecule. Enzymes that require a metal in their composition are known as METALLOENZYMES if they bind and retain their metal atom(s) under all conditions with very high affinity. While those which have a lower affinity for metal ion, but still require the metal ion for activity, are known as METAL-ACTIVATED ENZYMES. 134 Role of Coenzymes The functional role of coenzymes is to act as transporters of chemical groups from one reactant to another. The chemical groups carried can be as simple as the hydride ion (H+ + 2e-) carried by NAD or the mole of hydrogen carried by FAD; or they can be even more complex than the amine (-NH2) carried by pyridoxal phosphate. Since coenzymes are chemically changed as a consequence of enzyme action, it is often useful to consider coenzymes to be a special class of substrates, or second substrates, which are common to many different holoenzymes. In all cases, the coenzymes donate the carried chemical grouping to an acceptor molecule and are thus regenerated to their original form. This regeneration of coenzyme and holoenzyme fulfills the definition of an enzyme as a chemical catalyst, since (unlike the usual substrates, which are used up during the course of a reaction) coenzymes are generally regenerated. Enzyme Relative to Substrate Type Although enzymes are highly specific for the kind of reaction they catalyze, the same is not always true of substrates they attack. For example, Succinic dehydrogenase (SDH) always catalyzes an oxidation reduction reaction and its substrate is succinic acid. 135 Alcohol dehydrogenase (ADH) also catalyzes oxidation-reduction reactions but attacks a number of different alcohols, ranging from methanol to butanol. Generally, enzymes having broad substrate specificity are most active against one particular substrate. In the case of alcohol dehydrogenase, ethanol is the preferred substrate. Enzymes also are generally specific for a particular steric configuration (optical isomer) of a substrate. Enzymes that attack D-sugars will not attack the corresponding L- isomer. Enzymes that act on L-amino acids will not employ the corresponding D-optical isomer as a substrate. The enzymes known as racemases provide a striking exception to these generalities; in fact, the role of racemases is to convert D-isomers to Lisomers and vice versa. Thus racemases attack both D and L forms of their substrate. As enzymes have a more or less broad range of substrate specificity, it follows that a given substrate may be acted on by a number of different enzymes, each of which uses the same substrate(s) and produces the same product(s). The individual members of a set of enzymes sharing such characteristics are known as ISOZYMES. The best studied set of isozymes is the lactate dehydrogenase (LDH) system. 136 LDH is a tetrameric enzyme composed of all possible arrangements of two different protein subunits. These subunits combine in various combinations leading to 5 distinct isozymes. ENZYME-SUBSTRATE INTERACTIONS The favored model of enzyme substrate interaction is known as the induced fit model. This model proposes that the initial interaction between enzyme and substrate is relatively weak, but that these weak interactions rapidly induce conformational changes in the enzyme that strengthen binding and bring CATALYTIC SITES close to substrate bonds to be altered. Enzymes as Biological Catalysts In cells and organisms most reactions are catalyzed by enzymes, which are regenerated during the course of a reaction. These biological catalysts are physiologically important because they speed up the rates of reactions that would otherwise be too slow to support life. Enzymes increase reaction rates sometimes by as much as one million fold, but more typically by about one thousand fold. Catalysts speed up the forward and reverse reactions proportionately so that, although the magnitude of the rate constants of the forward and reverse reactions is are increased, the ratio of the rate constants remains the same in the presence or absence of enzyme. 137 Enzymes increase reaction rates by decreasing the amount of energy required to form a complex of reactants that is competent to produce reaction products. This complex is known as the activated state or transition state complex for the reaction. Michaelis-Menton Kinetics In typical enzyme-catalyzed reactions, reactant and product concentrations are usually hundreds or thousands of times greater than the enzyme concentration. Consequently, each enzyme molecule catalyzes the conversion to product of many reactant molecules. In biochemical reactions, reactants are commonly known as substrates. The catalytic event that converts substrate to product involves the formation of a transition state, and it occurs most easily at a specific binding site on the enzyme. This site, called the catalytic site of the enzyme, has been evolutionarily structured to provide specific, high-affinity binding of substrate(s) and to provide an environment that favors the catalytic events. The complex that forms when substrate(s) and enzyme combined, is called the enzyme substrate (ES) complex. Reaction products arise when the ES complex breaks down releasing free enzyme. Between the binding of substrate to enzyme, and the reappearance of free enzyme and product, a series of complex events must take place. At a minimum an ES complex must be formed; this complex must pass to the transition state (ES*); and the transition state complex must advance to an enzyme product complex (EP). The latter is finally 138 competent to dissociate to product and free enzyme. The series of events can be shown thus: E+S <--> EScomplex<--> ES*complex<--> EPcomplex<--> E + P The kinetics of simple reactions like that above were first characterized by biochemists Michaelis and Menten. The concepts underlying their analysis of enzyme kinetics continue to provide the cornerstone for understanding metabolism today, and for the development and clinical use of drugs aimed at selectively altering rate constants and interfering with the progress of disease states. The Michaelis-Menten equation is a quantitative description of the relationship among the rate of an enzyme- catalyzed reaction [v1], the concentration of substrate [S] and two constants, Vmax and Km (which are set by the particular equation). The symbols used in the Michaelis-Menton equation refer to the reaction rate [v1], maximum reaction rate (Vmax), substrate concentration [S] and the Michaelis-Menton constant (Km). The Michaelis-Menten equation can be used to demonstrate that at the substrate concentration that produces exactly half of the maximum reaction rate, i.e., 1/2 Vmax , the substrate concentration is numerically equal to Km. 139 This fact provides a simple yet powerful bioanalytical tool that has been used to characterize both normal and altered enzymes, such as those that produce the symptoms of genetic diseases. Rearranging the Michaelis-Menton equation leads to: From this equation it should be apparent that when the substrate concentration is half that required to support the maximum rate of reaction, the observed rate, v1, will, be equal to Vmax divided by 2; in other words, v1 = [Vmax/2]. At this substrate concentration Vmax/v1 will be exactly equal to 2, with the result that: Km =[S])2-1) = [S] The latter is an algebraic statement of the fact that, for enzymes of the Michaelis-Menten type, when the observed reaction rate is half of the maximum possible reaction rate, the substrate concentration (S) is numerically equal to the Michaelis-Menten constant (Km) . In this derivation, the units of Km are those used to specify the concentration of S, usually Molarity. 140 Plotting of substrate concentration versus reaction velocity in Michaelis-Menten equation: The key features of the plot are marked by points A, B and C. At high substrate concentrations the rate represented by point C the rate of the reaction is almost equal to Vmax, and the difference in rate at nearby concentrations of substrate is almost negligible. If the Michaelis-Menten plot is extrapolated to infinitely high substrate concentrations, the extrapolated rate is equal to Vmax. When the reaction rate becomes independent of substrate concentration, or nearly so, the rate is said to be zero order. 141 The very small differences in reaction velocity at substrate concentrations around point C (near Vmax) reflect the fact that at these concentrations almost all of the enzyme molecules are bound to substrate and the rate is virtually independent of substrate, hence zero order. At lower substrate concentrations, such as at points A and B, the lower reaction velocities indicate that at any moment only a portion of the enzyme molecules are bound to the substrate. In fact, at the substrate concentration denoted by point B, exactly half the enzyme molecules are in an EScomplex at any instant and the rate is exactly one half of Vmax. At substrate concentrations near point A the rate appears to be directly proportional to substrate concentration, and the reaction rate is said to be first order. Inhibition of Enzyme Catalyzed Reactions To avoid dealing with curvilinear plots of enzyme catalyzed reactions, biochemists Lineweaver and Burk introduced an analysis of enzyme kinetics based on the following rearrangement of the Michaelis-Menten equation: Take the inverse: 1/v1 = Km /Vmax[S] + 1/Vmax 142 Plots of 1/v1 versus 1/[S] yield straight lines having a slope of Km/Vmax and an intercept on the ordinate at 1/Vmax. A Lineweaver-Burk Plot Lineweaver-Burk transformation of the Michaelis-Menton equation is useful in the analysis of enzyme inhibition. Since most clinical drug therapy is based on inhibiting the activity of enzymes, analysis of enzyme reactions using the tools described above has been fundamental to the modern design of pharmaceuticals. Enzyme inhibitors fall into two broad classes: 1. Inhibitors causing irreversible inactivation of enzymes. 2. Inhibitors whose inhibitory effects can be reversed. 3. Irreversible Inhibitors: 143 They cause an inactivating, covalent modification of enzyme structure. Cyanide is a classic example of an irreversible enzyme inhibitor by covalently binding mitochondrial cytochrome oxidase, it inhibits all the reactions associated with electron transport. The kinetic effect of irreversible inhibitors is to decrease the concentration of active enzyme, thus decreasing the maximum possible concentration of EScomplex . Since the limiting enzyme reaction rate is often k2[ES], it is clear that under these circumstances the reduction of enzyme concentration will lead to decreased reaction rates. Note that when enzymes in cells are only partially inhibited by irreversible inhibitors, the remaining unmodified enzyme molecules are not distinguishable from those in untreated cells; in particular, they have the same turnover number and the same Km. Turnover number, related to Vmax, is defined as the maximum number of moles of substrate that can be converted to product per mole of catalytic site per second. Irreversible inhibitors are usually considered to be poisons and are generally unsuitable for therapeutic purposes. Reversible inhibitors: They can be divided into three categories: 1. Competitive inhibitors. 2. Noncompetitive inhibitors. 3. Uncompetitive inhibitors. 144 Inhibitor Type Binding Site on Enzyme Kinetic effect Specifically at the catalytic site. Competitive Inhibitor It competes with substrate Vmax is unchanged. for binding. Km Km appears unaltered. Vmax is increased. Inhibition is reversible by substrate. Binds E or ES complex other than at the catalytic site. Noncompetitive Inhibitor Substrate binding is decreased unaltered, but ESI complex proportionately cannot form products. inhibitor conc. Inhibition cannot to be reversed by substrate. Binds only to ES complexes at locations other than the catalytic site. Uncompetitive Substrate binding modifies enzyme structure, making Inhibitor inhibitor- binding site cannot be available. Inhibition reversed by substrate. 145 Apparent decreased. Km is decreased. Vmax When the reversible inhibitor concentration drops, enzyme activity is regenerated. Usually these inhibitors bind to enzymes by non-covalent forces and the inhibitor maintains a reversible equilibrium with the enzyme. The equilibrium constant for the dissociation of enzyme inhibitor complexes is known as KI: KI = [E] [I] / [EI complex] The importance of KI is that in all enzyme reactions where substrate, inhibitor and enzyme interact, the normal Km and or Vmax for substrate enzyme interaction appear to be altered. These changes are a consequence of the influence of KI on the overall rate equation for the reaction. The effects of KI are best observed in Lineweaver-Burk plots. Probably the best known reversible inhibitors are competitive inhibitors, which always bind at the catalytic or active site of the enzyme. Most drugs that alter enzyme activity are of this type. Competitive inhibitors are especially attractive as clinical modulators of enzyme activity because they offer two routes for the reversal of enzyme inhibition, while other reversible inhibitors offer only one. First, as with all kinds of reversible inhibitors, a decreasing concentration of the inhibitor reverses the equilibrium regenerating active free enzyme. Second, since substrate and competitive inhibitors both bind at the same site they compete with one another for binding 146 Raising the concentration of substrate (S), while holding the concentration of inhibitor constant, provides the second route for reversal of competitive inhibition. The greater the proportion of substrate, the greater the proportion of enzyme present in competent ES complexes. As noted earlier, high concentrations of substrate can displace virtually all competitive inhibitor bound to active sites. Thus, it is apparent that Vmax should be unchanged by competitive inhibitors. 147 Lineweaver-Burk Plots of Inhibited Enzymes 148 4. NUCLEIC ACID Nucleic acids were found to be major components of chromosomes, small gene-carrying bodies in the nuclei of complex cells. Elemental analysis of nucleic acids showed the presence of phosphorus, in addition to the usual C, H, N & O. Unlike proteins, nucleic acids contained no sulfur. Complete hydrolysis of chromosomal nucleic acids gave inorganic phosphate, 2-deoxyribose (a previously unknown sugar) and four different heterocyclic bases (shown in the following diagram). To reflect the unusual sugar component, chromosomal nucleic acids are called deoxyribonucleic acids, abbreviated DNA. Analogous nucleic acids in which the sugar component is ribose are termed ribonucleic acids, abbreviated RNA. The acidic character of the nucleic acids was attributed to the phosphoric acid moiety. 149 Chemical structure The term "nucleic acid" is the generic name for a family of biopolymers, named for their role in the cell nucleus. The monomers from which nucleic acids are constructed are called nucleotides. Each nucleotide consists of three components: Nitrogenous heterocyclic base, which is either a purine or a pyrimidine. Pentose sugar. Phosphate group. Nucleic acid types differ in the structure of the sugar in their nucleotides - DNA contains 2-deoxyriboses while RNA contains ribose (where the only difference is the presence of a hydroxyl group). Also, the nitrogenous bases found in the two nucleic acid types are different: adenine, cytosine, and guanine are found in both RNA and DNA, while thymine only occurs in DNA and uracil only occurs in RNA. Types of nucleic acids Ribonucleic acid Ribonucleic acid, or RNA, is a nucleic acid polymer consisting of nucleotide monomers, which plays several important roles in the processes of translating genetic information from deoxyribonucleic acid (DNA) into proteins. RNA acts as a messenger between DNA and the protein synthesis complexes known as ribosomes, forms vital portions of ribosomes, and serves as an essential carrier molecule for amino acids to be used in protein synthesis. 150 Deoxyribonucleic acid Deoxyribonucleic acid (DNA) is a nucleic acid that contains the genetic instructions used in the development and functioning of all known living organisms. The main role of DNA molecules is the long-term storage of information and DNA is often compared to a set of blueprints, since it contains the instructions needed to construct other components of cells, such as proteins and RNA molecules. The DNA segments that carry this genetic information are called genes, but other DNA sequences have structural purposes, or are involved in regulating the use of this genetic information. 151 The two monocyclic bases shown here are classified as pyrimidines, and the two bicyclic bases are purines. Each has at least one N-H site at which an organic substituent may be attached. They are all polyfunctional bases, and may exist in tautomeric forms. The Chemical Nature of DNA The polymeric structure of DNA may be described in terms of monomeric units of increasing complexity. Condensation polymerization of these leads to the DNA formulation outlined above. Finally, a 5'- monophosphate ester, called a nucleotide may be drawn as a single monomer unit. Since a monophosphate ester of this kind is a strong acid, it will be fully ionized at the usual physiological pH (ca.7.4). DNA components are given by selective hydrolysis of DNA through the action of nuclease enzymes. Anhydride-like di- and triphosphate nucleotides have been identified as important energy carriers in biochemical reactions, the most common being ATP (adenosine 5'-triphosphate). 152 153 A complete structural representation of a segment of the DNA polymer formed from 5'-nucleotides. Several important characteristics of this formula should be noted. • First, the remaining P-OH function is quite acidic and is completely ionized in biological systems. • Second, the polymer chain is structurally directed. One end (5') is different from the other (3'). • Third, although this appears to be a relatively simple polymer, the possible permutations of the four nucleosides in the chain become very large as the chain lengthens. • Fourth, the DNA polymer is much larger than originally believed. Molecular weights for the DNA from multicellular organisms are commonly 109 or greater. Nucleic acid components Nucleobases Nucleobases are heterocyclic aromatic organic compounds containing nitrogen atoms. Nucleobases are the parts of RNA and DNA involved in base pairing. Cytosine, guanine, adenine, thymine are found predominantly in DNA, while in RNA uracil replaces thymine. These are abbreviated as C, G, A, T, U, respectively. Nucleobases are complementary, and when forming base pairs, must always join accordingly: cytosine-guanine, adenine-thymine (adenineuracil when RNA). The strength of the interaction between cytosine and guanine is stronger than between adenine and thymine because the former pair has three hydrogen bonds joining them while the latter pair have only two. Thus, the higher the GC content of double-stranded DNA, the more stable the molecule and the higher the melting temperature. 154 Two main nucleobase classes exist, named for the molecule which forms their skeleton. These are the double-ringed purines and singleringed pyrimidines. Adenine and guanine are purines (abbreviated as R), while cytosine, thymine, and uracil are all pyrimidines (abbreviated as Y). Hypoxanthine and xanthine are mutant forms of adenine and guanine, respectively, created through mutagen presence, through deamination (replacement of the amine-group with a hydroxyl-group). These are abbreviated HX and X. Nucleosides Nucleosides are glycosylamines made by attaching a nucleobase (often referred to simply as bases) to a ribose or deoxyribose (sugar) ring. In short, a nucleoside is a base linked to sugar. The names derive from the nucleobase names. The nucleosides commonly occurring in DNA and RNA include cytidine, uridine, adenosine, guanosine and thymidine. When a phosphate is added to a nucleoside (by phosphorylated by a specific kinase enzyme), a nucleotide is produced. Nucleoside analogues, such as acyclovir, are used as antiviral agents. Nucleotides and deoxynucleotides A nucleotide consists of a nucleoside and one or more phosphate groups. Nucleotides are the monomers of RNA and DNA, as well as forming the structural units of several important cofactors - CoA, flavin adenine dinucleotide, flavin mononucleotide, adenosine triphosphate and nicotinamide adenine dinucleotide phosphate. In the cell nucleotides play important roles in metabolism, and signaling. Nucleotides are named after the nucleoside on which they are based, in conjunction with the number of phosphates they contain, for example: 155 Adenine bonded to ribose forms the nucleoside adenosine. Adenosine bonded to a phosphate forms adenosine monophosphate. As phosphates are added, adenosine diphosphate and adenosine triphosphate are formed, in sequence. 156 5. VITAMINS & MINERALS Vitamins are organic substances (made by plants or animals), minerals are inorganic elements that come from the earth; soil and water and are absorbed by plants. Animals and humans absorb minerals from the plants they eat. Vitamins and minerals are nutrients that your body needs to grow and develop normally. Vitamins and minerals, have a unique role to play in maintaining your health. For example Vitamin D helps your body absorb the amount of calcium (a mineral) it needs to form strong bones. A deficiency in vitamin D can result in a disease called rickets (softening of the bones caused by the bodies inability to absorb the mineral calcium.) The body cannot produce calcium; therefore, it must be absorbed through our food. Other minerals like chromium, copper, iodine, iron, selenium, and zinc are called trace minerals because you only need very small amounts of them each day. The best way to get enough vitamins is to eat a balanced diet with a variety of foods. You can usually get all your vitamins from the foods you eat. 157 Vitamins Hang Out in Water and Fat There are two types of vitamins: fat soluble and water soluble. When you eat foods that contain fat-soluble vitamins, the vitamins are stored in the fat tissues in your body and in your liver. They wait around in your body fat until your body needs them. Fat-soluble vitamins are happy to stay stored in your body for awhile — some stay for a few days, some for up to 6 months! Then, when it's time for them to be used, special carriers in your body take them to where they're needed. Vitamins A, D, E, and K are all fat-soluble vitamins. Water-soluble vitamins are different. When you eat foods that have water-soluble vitamins, the vitamins don't get stored as much in your body. Instead, they travel through your bloodstream. Whatever your body doesn't use comes out when you urinate (pee). So these kinds of vitamins need to be replaced often because they don't stick around! This crowd of vitamins includes vitamin C and the big group of B vitamins — B1 (thiamin), B2 (riboflavin), niacin, B6 (pyridoxine), folic acid, B12 (cobalamine), biotin, and pantothenic acid. Vitamins Feed Your Needs Your body is one powerful machine, capable of doing all sorts of things by itself. But one thing it can't do is make vitamins. That's where food comes in. Your body is able to get the vitamins it needs from the foods you eat because different foods contain different vitamins. The key is to eat different foods to get an assortment of vitamins. Though some kids take a daily vitamin, most kids don't need one if they're eating a variety of healthy foods. 158 VITAMINS & COENZYMES Vitamins are organic molecules that function in a wide variety of capacities within the body. The most prominent function is as cofactors for enzymatic reactions. They generally cannot be synthesized by mammalian cells and therefore, must be supplied in the diet. The vitamins are of two types: 1. Water Soluble Vitamins 2. Fat Soluble Vitamins Thiamin (B1) Riboflavin (B2) Niacin (B3) Vitamin A Pantothenic Acid (B5) Vitamin D Pyridoxine (B6) Vitamin E Biotin Vitamin K Cobalamin (B12) Folic Acid Ascorbic Acid 159 Thiamin (Vitamin B1) Thiamin is also known as vitamin B1 . Thiamin is rapidly converted to its active form, thiamin pyrophosphate, TPP, in the brain and liver by a specific enzymes, thiamin diphosphotransferase. Thiamin pyrophosphate (TPP) 160 TPP is necessary as a cofactor for the pyruvate and ketoglutarate dehydrogenase catalyzed reactions as well as the transketolase catalyzed reactions of the pentose phosphate pathway. The dietary requirement for thiamin is proportional to the caloric intake of the diet and ranges from 1.0 - 1.5 mg/day for normal adults. The severe thiamin deficiency disease known as Beriberi مرض الهزال والضعف, is the result of a diet that is carbohydrate rich and thiamin deficient. Riboflavin (Vitamin B2) Riboflavin is the precursor for the coenzymes, flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD). The enzymes that require FMN or FAD as cofactors are termed flavoproteins. Several flavoproteins also contain metal ions and are termed metalloflavoproteins. 161 Both classes of enzymes flavoproteins and metalloflavoproteins are involved in a wide range of redox reactions, e.g. succinate dehydrogenase and xanthine oxidase. During the course of the enzymatic reactions involving the flavoproteins, the reduced forms of FMN and FAD are formed, FMNH2 and FADH2, respectively. The normal daily requirement for riboflavin is 1.2 - 1.7 mg/day for normal adults. Structure of nitrogens 1 & 5 carry hydrogens in FADH2 FAD Riboflavin is present in eggs, milk, meat and cereals. Riboflavin decomposes when exposed to visible light. This characteristic can lead to riboflavin deficiencies in newborns treated 162 for hyperbilirubinemia (Increase in bilirubin in blood, bilirubin is a bile pigment which is a degradation product of heme)) by phototherapy. Vitamin B3 Niacin (nicotinamide & nicotinic acid) Nicotinamide Nicotinic Acid Niacin (nicotinic acid and nicotinamide) is also known as vitamin B3. Both nicotinic acid and nicotinamide can serve as the dietary source of vitamin B3. Niacin is required for the synthesis of the active forms of vitamin B 3, nicotinamide adenine dinucleotide (NAD+) and nicotinamide adenine dinucleotide phosphate (NADP+). Both NAD+ and NADP+ function as cofactors for numerous dehydrogenase, e.g., lactate and malate dehydrogenases. 163 Structure NADH of is shown in NAD+ the box insert. The -OH phosphorylated in NADP+ is indicated by the red arrow. Niacin is not a true vitamin in the strictest definition since it can be derived from the amino acid tryptophan. Also, synthesis of niacin from tryptophan requires vitamins B1, B2 and B6 . The recommended daily requirement for niacin is 13 - 19 niacin equivalents (NE) per day for a normal adult. One NE is equivalent to 1 mg of free niacin). The severe symptoms, depression, dermatitis and diarrhea, are associated with the condition known as pellagra )داء ( مرض جلدي البالجرا 164 Nicotinic acid (but not nicotinamide) when administered in pharmacological doses of 2 - 4 g/day lowers plasma cholesterol levels and has been shown to be a useful therapeutic for hypercholesterolemia. The major action of nicotinic acid in this capacity is a reduction in fatty acid mobilization from adipose tissue. Although nicotinic acid therapy lowers blood cholesterol it also causes a depletion of glycogen stores and fat reserves in skeletal and cardiac muscle. Additionally, there is an elevation in blood glucose and uric acid production. Pantothenic Acid (Vitamin B5) Pantothenic acid is formed from alanine and pantoic acid. Pantothenate is required for synthesis of coenzyme A, CoA and is a component of the acyl carrier protein (ACP) domain of fatty acid synthase. 165 Coenzyme A Pantothenate is, therefore, required for the metabolism of carbohydrate via the TCA cycle and all fats and proteins. At least 70 enzymes have been identified as requiring CoA or ACP derivatives for their function. Deficiency of pantothenic acid is extremely rare due to its widespread distribution in whole grain cereals, legumes and meat. Symptoms of pantothenate deficiency are difficult to assess since they are subtle and resemble those of other B vitamin deficiencies. 166 Vitamin B6 Pyridoxine Pyridoxal Pyridoxamine Pyridoxal, pyridoxamine and pyridoxine are collectively known as vitamin B6. All three compounds are efficiently converted to the biologically active form of vitamin B6, pyridoxal phosphate. This conversion is catalyzed by the ATP requiring enzyme, pyridoxal kinase. Pyridoxal Phosphate Pyridoxal phosphate functions as a cofactor in enzymes involved in transamination reactions required for the synthesis and catabolism of the amino acids as well as in glycogenolysis as a cofactor for glycogen phosphorylase. The requirement for vitamin B6 in the diet is proportional to the level of protein consumption ranging from 1.4 - 2.0 167 mg/day for a normal adult. During pregnancy and lactation the requirement for vitamin B6 increases approximately 0.6 mg/day. Deficiencies of vitamin B6 are rare and usually are related to an overall deficiency of all the B-complex vitamins. Biotin Biotin is the cofactor required of enzymes that are involved in carboxylation reactions, e.g. acetyl-CoA carboxylase and pyruvate carboxylase. Biotin is found in numerous foods and also is synthesized by intestinal bacteria and as such deficiencies of the vitamin are rare. Deficiencies are generally seen only after long antibiotic therapies which deplete the intestinal fauna or following excessive consumption of raw eggs. The latter is due to the affinity of the egg white protein, avidin, for biotin preventing intestinal absorption of the biotin. 168 Cobalamin (Vitamin B12) Vitamin B12 is composed of a complex tetrapyrrol ring structure (corrin ring) and a cobalt ion in the center. Vitamin B12 is synthesized exclusively by microorganisms and is found in the liver of animals bound to protein as methycobalamin or 5'-deoxy adenosyl cobalamin. The vitamin must be hydrolyzed from protein in order to be active. Hydrolysis occurs in the stomach by gastric acids or the intestines by trypsin digestion following consumption of animal meat. The vitamin is then bound by intrinsic factor, a protein secreted by parietal cells الخاليا الجداريةof the stomach, and carried to the ileum المعي اللفائفيwhere it is absorbed. Following absorption the vitamin is transported to the liver in the blood bound to transcobalamin II. There are only two clinically significant reactions in the body that require vitamin B12 as a cofactor: 1. During the conversion of methylmalonyl-CoA to succinyl-CoA. 2. For catalysis of the conversion of homocysteine to methionine. 169 Cyanocobalamin Pernicious anemia فقر الدم الخبيثis a megaloblastic anemia resulting from vitamin B12 deficiency that develops as a result a lack of intrinsic factor in the stomach leading to malabsorption of the vitamin. 170 Folic Acid Folic Acid Folic acid is obtained primarily from yeasts and leafy vegetables as well as animal liver. Folic acid is reduced within cells (principally the liver where it is stored) to tetrahydrofolate (THF). The function of THF derivatives is to carry and transfer various forms of one carbon units during biosynthetic reactions. The one carbon units are either methyl, methylene, methenyl, formyl or formimino groups. 171 These one carbon transfer reactions are required in the biosynthesis of serine, methionine, glycine, choline and the purine nucleotides and dTMP. The most pronounced effect of folate deficiency on cellular processes is upon DNA synthesis. The result is MEGALOBLASTIC ANEMIA as for vitamin B12 deficiency. The inability to synthesize DNA during erythrocyte maturation leads to abnormally large erythrocytes termed MACROCYTIC ANEMIA. 172 Ascorbic Acid (Vitamin C) Ascorbic acid is derived from glucose via the uronic acid pathway. The main function of ascorbate is as a reducing agent in a number of different reactions. Vitamin C has the potential to reduce cytochromes A and C of the respiratory chain as well as molecular oxygen. The most important reaction requiring ascorbate as a cofactor is the hydroxylation of proline residues in collagen. Vitamin C is, therefore, required for the maintenance of normal connective tissue as well as for wound healing since synthesis of connective tissue is the first event in wound tissue remodeling. Vitamin C also is necessary for bone remodeling due to the presence of collagen in the organic matrix of bones. Several other metabolic reactions require vitamin C as a cofactor. These include the catabolism of tyrosine and the synthesis of epinephrine from tyrosine and the synthesis of the bile acids. It is also believed that vitamin C is involved in the process of steroidogenesis since the adrenal cortex contains high levels of vitamin C which are depleted upon adrenocorticotropic hormone (ACTH) stimulation of the gland. 173 The disease SCURVY ) (داء االسقربوط تورم اللثة ونزف الدم منهاdue to the role of the vitamin in the post-translational modification of collagens. المادة البروتينية في النسيج والعظم Vitamin C is readily absorbed and so the primary cause of vitamin C deficiency is poor diet and/or an increased requirement. Soluble vitamins in lipid Vitamin A Vitamin A consists of three biologically active molecules, retinol, retinal and retinoic acid. Retinal Retinol Retinoic Acid 174 Each of these compounds is derived from the plant precursor molecule, β-carotene (a member of a family of molecules known as carotenoids). β-carotene, which consists of two molecules of retinal linked at their aldehyde ends, is also referred to as the provitamin form of vitamin A. Ingested β -carotene is cleaved in the lumen تجويفof the intestine by β -carotene dioxygenase to yield retinal. Retinal is reduced to retinol. Retinol is esterified to palmitic acid and delivered to the blood. The earliest symptoms of vitamin A deficiency are night blindness. Prolonged lack of vitamin A leads to deterioration of the eye tissue through progressive keratinization of the cornea, a condition known as xerophthalmia. جفاف العين Beta-carotene is a very effective antioxidant and is suspected to reduce the risk of cancers known to be initiated by the production of free radicals. Of particular interest is the potential benefit of increased β-carotene intake to reduce the risk of lung cancer in smokers. However, caution needs to be taken when increasing the intake of any of the lipid soluble vitamins. Excess accumulation of vitamin A in the liver can lead to toxicity which manifests as bone pain, hepatosplenomegaly, nausea and diarrhea. 175 Vitamin D Ergosterol 7-Dehydrocholesterol Vitamin D2 Vitamin D3 Vitamin D is a steroid hormone that functions to regulate specific gene expression following interaction with its intracellular receptor. 176 The biologically active form of the hormone is 1,25-dihydroxy vitamin D3 (1,25-(OH)2D3, also termed calcitriol). Calcitriol functions primarily to regulate calcium and phosphorous homeostasis. Ergocalciferol (vitamin D2) is formed by uv irradiation of ergosterol. In the skin 7-dehydrocholesterol is converted to cholecalciferol (vitamin D3) following uv irradiation. Vitamin D2 and D3 are processed to D2-calcitriol and D3-calcitriol, respectively, by the same enzymatic pathways in the body. The main symptom of vitamin D deficiency in children is rickets داء كساح األطفالand in adults is osteomalacia لين العظام. Rickets is characterized improper mineralization development of the bones resulting in soft bones. Vitamin E Tocopherol 177 during the Vitamin E is a mixture of several related compounds known as tocopherols. Due to its Lipophilic nature, vitamin E accumulates in cellular membranes, fat deposits and other circulating lipoproteins. The major site of vitamin E storage is in adipose tissue. The major function of vitamin E is to act as a natural antioxidant by scavenging free radicals and molecular oxygen. In particular vitamin E is important for preventing peroxidation of polyunsaturated membrane fatty acids. The vitamins E and C are interrelated in their antioxidant capabilities. Active tocopherol can be regenerated by interaction with vitamin C following scavenge of a peroxy free radical. Alternatively, tocopherol can scavenge two peroxy free radicals and then be conjugated to glucuronate for excretion in the bile. The major symptom of vitamin E deficiency in humans is an increase in red blood cell fragility قابلية التكسير. Since vitamin E is absorbed from the intestines, any fat malabsorption diseases can lead to deficiencies in vitamin E intake. Vitamin K The K vitamins exist naturally as K1 (phylloquinone) in green vegetables and K2 (menaquinone) produced by intestinal bacteria and K3 is synthetic menadione. 178 When administered, vitamin K3 is alkylated to one of the vitamin K2 forms of menaquinone. Vitamin K1 Vitamin K2 Vitamin K3 The major function of the K vitamins is in the maintenance of normal levels of the blood clotting proteins. Fat malabsorptive diseases can result in vitamin K deficiency. The primary symptom of a deficiency in infants is a hemorrhagic syndrome. 179 Minerals Minerals are inorganic micronutrients. They are categorized into 2 groups according to their amounts in our bodies. Major minerals include calcium, chloride, magnesium, phosphorus, potassium; sodium and sulphur, are found in our bodies in amounts greater than 5 grams. However, trace minerals which are found in an amount less than 5 grams include chromium, copper, fluoride, iodine, iron, manganese, selenium and zinc. CALCIUM: Builds bones and teeth; aids in proper function of muscles, heart, nerves, and iron utilization; helps blood coagulation; regulates the passage of nutrients in and out of cells; relieves pain and cramps; eases insomnia. NATURAL SOURCES: Dairy products, soybeans, sunflower seeds, legumes, sardines. MAGNESIUM: Reduces blood cholesterol; forms hard tooth enamel and fights tooth decay; aids in converting blood sugar into energy; helps regulate body temperature; aids nerve function and bone growth; helps utilize Vitamins B, C, E; promotes absorption and metabolism of other minerals; activates enzymes for metabolism of carbohydrates and amino acids; prevents calcium deposits in the bladder, heart 180 attacks, depression, polio. NATURAL RESOURCES: Nuts, figs, seeds, dark-green vegetables, wheat bran, avocados, bananas. IRON: Present in all cells; one of the constituents of hemoglobin which carries oxygen to the tissues by blood circulation. NATURAL RESOURCES: Liver, meat, raw clams, oysters, oatmeal, nuts, beans, wheat germ. IODINE: Aids thyroid gland and prevents goiter; helps burn fat; converts carotene into Vitamin A; aids absorption of carbohydrates from small intestine; promotes growth; regulates energy production; maintains hair, nails skin and teeth. NATURAL SOURCES: Kelp, seafood, vegetables. Vitamins and minerals Vitamin or Mineral Examples of Good Food Sources What It Does Recommended Daily Amount (RDA) or Adequate Upper Limit (The Highest Amount You Can Take Without Risk Calcium Milk, yogurt, Essential for bone hard cheeses, growth and fortified cereals, strength, blood spinach clotting, muscle contraction, and the transmission of nerve signals Adults age 19-50: 1,000 milligrams/day Adults age 51 and up: 1,200 milligrams/day 2,500 milligrams/day Choline (Vitamin B complex) Milk, liver, eggs, Plays a key role in peanuts the production of cells and neurotransmitters Men: 550 milligrams/day Women: 425 milligrams/day Pregnantwomen: 450 milligrams/day Breastfeedingwomen: 550 milligrams/day 3,500 milligrams/day Chromium Meats, poultry, fish, some cereals Helps controlblood sugar levels Adult men age 19-50: 35 micrograms/day Adult men age 51 and up: 30 micrograms/day Adult women age 19-50: 25 micrograms/day Adult women age 51 and up:20 181 Unknown micrograms/day Pregnant women: 30 micrograms/day Breastfeeding women: 45 micrograms/day Copper Seafood, nuts, seeds, wheat bran cereals, whole grains Fiber Bran cereal, peas, lentils, black beans, fruits, vegetables Fluoride Important in Adults: 900 themetabolism of micrograms/dayPregnant iron women: 1,000 micrograms/dayBreastfeeding women: 1,300 micrograms/day Helps with digestion and the maintenance of blood sugar levels; reduces the risk of heartdisease Adult men age 19-50: 38 grams/dayAdult men age 51 and up: 30 grams/dayAdult women age 19-50: 25 grams/dayAdult women age 51 and up:21 grams/dayPregnant women: 28 grams/dayBreastfeeding women: 29 grams/day Fluoridated Prevents the Adult men: 4 milligrams/dayAdult water, some sea formation women (including pregnant and fish, oftoothcavities an breastfeeding):3 milligrams/day sometoothpaste d stimulates the sand mouth growth of bone rinses 10,000 micrograms/day None 10 milligrams/day Folic Acid (Folate) Dark, leafy vegetables; enriched and whole grain breads; fortified cereals Key for the development of cells, protein metabolism andheart health; in pregnant women, helps prevent birth defects Adults: 400 micrograms/dayPregnant women: 600 micrograms/dayBreastfeeding women: 500 micrograms/day 1,000 micrograms/day Iodine Processed foods and iodized salt Important in the production ofthyroid hormon es Adults: 150 micrograms/day Pregnant women: 220 micrograms/day Breastfeeding women: 290 micrograms/day 1,100 micrograms/day Iron Magnesium Fortified cereals, Key component of beans, lentils, red blood cells beef, eggs and many enzymes Green leafy vegetables, Brazil nuts, almonds, soybeans, halibut, quinoa Helps with heart rhythm, muscle and nerve function, bone strength Men: 8 milligrams/dayWomen age 19-50: 18 milligrams/dayWomen age 51 and up: 8 milligrams/dayPregnant women: 27 milligrams/day Breastfeeding women: 9 milligrams/day 45 milligrams/day Adult men age 19-30: 400 milligrams/dayAdult men age 31 and up: 420 milligrams/dayAdult women age 19-30: 310 milligrams/day Adult women age 31 and up: 320 For magnesium in food and water, there is no upper limit. 182 For magnesium milligrams/dayPregnant women: 350-360 milligrams/dayBreastfeeding women: 310-320 milligrams/day in supplementsor fortified foods: 350 milligrams/day Manganese Nuts, beans and other legumes, tea, whole grains Important in forming bones and some enzymes Men: 2.3 milligrams/day Adult women: 1.8 milligrams/day Pregnant women: 2.0 milligrams/day Breastfeeding women: 2.6 milligrams/day 11 milligrams/day Molybdenum Legumes, grains, nuts Key in the production of some enzymes Adults: 45 micrograms/dayPregnant and breastfeeding women: 50 micrograms/day 2,000 micrograms/day Phosphorus Potassium Milk and other Allows cells to Adults: 700 milligrams/day dairy products, function normally; peas, meat, helps the body eggs, some produce energy; cereals and key in bone breads growth Sweet potato, bananas, yogurt, yellowfin tuna, soybeans Important in Adults: 4,700 milligrams per maintaining dayBreastfeeding women: 5,100 normal fluid milligrams/day balance; helps control blood pressure; reduces risk of kidney stones Selenium Organ meats, Protects cells seafood, some from damage; plants (if grown regulates thyroid in soil with hormone selenium) Brazil nuts. Adults: 55 micrograms/dayPregnant women: 60 micrograms/dayBreastfeeding women: 70 micrograms/day Sodium Foods to which Important for fluid sodium chloride balance (salt) has been added, like salted meats, nuts, butter, and a vast number of processed foods Adults age 19-50: 1500 milligrams/dayAdults age 5170: 1,300 milligrams/dayAdults age 71 and up: 1,200 milligrams/day Vitamin A Sweet potato Necessary for Men: 900 with peel, normal vision, micrograms/dayWomen: 700 carrots, spinach, immune function, micrograms/day fortified cereals reproduction 183 Adults up to age 70: 4,000 milligrams/dayAd ults over age 70: 3,000 milligrams/dayPre gnant women: 3500 milligrams/dayBre astfeeding women: 4,000 milligrams/day Unknown 400 micrograms/day 2,300 milligrams/day 3,000 micrograms/day Vitamin B1(Thiamin) Whole grain, Allows the body enriched, to process fortified carbohydrates products; bread; and some protein. cereals Men: 1.2 milligrams/dayWomen: 1.1 milligrams/dayPregnant and breastfeeding women: 1.4 milligrams/day Unknown Vitamin B2(Riboflavin) Milk, bread Key in metabolism products, and the fortified cereals conversion of food into energy; helps produce red blood cells Men: 1.3 milligrams/day Women: 1.1 milligrams/day Pregnant Women: 1.4 milligrams/day Breastfeeding Women: 1.6 milligrams/day Unknown Vitamin B3(Niacin) Vitamin B5(Pantothenic Acid) Vitamin B6 Meat, fish, Assists in Men: 16 milligrams/dayWomen: 14 poultry, enriched digestion and the milligrams/dayPregnant and whole grain conversion of Women: 18 milligrams/day breads, fortified food into energy; ?Breastfeeding women: 17 cereals important in the milligrams/day production of cholesterol Chicken, beef, potatoes, oats, cereals, tomatoes Important in fatty Adults: 5 milligrams/dayPregnant acid metabolism women: 6 milligrams/dayBreastfeeding women: 7 milligrams/day Fortified cereals, Important for the fortified soy nervous system; products, organ helps the body meats metabolize proteins and sugar Men age 19-50:1.3 milligrams/dayMen age 51 up: 1.7 milligrams/dayWomen age 1950: 1.3 milligrams/dayWomen age 51 up: 1.5 milligrams/dayPregnant women: 1.9 milligrams/dayBreastfeeding women: 2 milligrams/day For niacin in natural sources, there is no upper limit. For niacin in supplements or fortified foods: 35 milligrams/day Unknown 100 milligrams/day Vitamin B7(Biotin) Liver, fruits, meats Helps with the Adults: 30 synthesis of fats, micrograms/dayBreastfeeding glycogen and women: 35 micrograms/day amino acids Unknown Vitamin B12(Cobalamin) Fish, poultry, meat, fortified cereals Important in the Adults: 2.4 production of red micrograms/dayPregnant blood cells women: 2.6 micrograms/dayBreastfeeding women: 2.8 micrograms/day Unknown Vitamin C Red and green peppers, kiwis, oranges, strawberries, broccoli Antioxidant that protects against cell damage, boosts the immune system, forms collagen in the body Men: 90 milligrams/dayWomen: 75 milligrams/dayPregnant women: 85 milligrams/dayBreastfeeding women: 120 milligrams/day 2,000 milligrams/day Vitamin D (Calciferol) Fish liver oils, fatty fish, fortified milk Crucial in metabolizing calcium for Adults age 18-50: 5 micrograms/dayAdults age 5170: 10 micrograms/dayAdults over 50 micrograms/day 184 products, fortified cereals; also, formed naturally as a result of sunlight exposure Vitamin E (alphatocopherol) Vitamin K Zinc Fortified cereals, sunflower seeds, almonds, peanut butter, vegetable oils healthy bones age 70: 15 micrograms/dayPregnant and breastfeeding women: 5 micrograms/day Antioxidant that Adults (including pregnant protects cells women): 15 against damage milligrams/dayBreastfeeding women: 19 Green Important in blood Men: 120 micrograms/day vegetables like clotting and bone ___ spinach, health Women (including pregnant and collards, and breastfeeding):90 micrograms/day broccoli; brussels sprouts; cabbage Red meats, some seafood, fortified cereals Supports the body's immunity and nerve function; important in reproduction Men: 11 milligrams/day ___ Women: 8 milligrams/dayPregnant women: 11 milligrams/dayBreastfeeding women: 12 milligrams/day 185 1,000 milligrams/day Unknown 40 milligrams/day 6.HORMONE A hormone is a chemical released by one or more cells that affects cells in other parts of the organism. Only a small amount of hormone is required to alter cell metabolism. Hormone is essentially a chemical messenger that transports a signal from one cell to another. Hormones in animals are often transported in the blood. Cells respond to a hormone when they express a specific receptor for that hormone. The hormone binds to the receptor protein, resulting in the activation of a signal transduction mechanism that ultimately leads to cell typespecific responses. Endocrine hormone molecules are secreted (released) directly into the bloodstream. Exocrine hormone (or ecto-hormones) are secreted directly into a duct, and from the duct they either flow into the bloodstream or they flow from cell to cell by diffusion in a process known as paracrine signalling. Chemical classes of hormones There are three different classes of hormone based on their chemical composition: 1. Amines Amines, such as nor-epinephrine, epinephrine, and dopamine, are derived from single amino acids, in this case tyrosine. 186 Thyroid hormones such as 3,5,3’-tri-iodothyronine (T3) and 3,5,3’,5’tetra-iodothyronine (thyroxine, T4) make up a subset of this class because they derive from the combination of two iodinated tyrosine amino acid residues. 2. Peptide and protein Peptide hormones and protein hormones consist of three (in the case of thyrotropin-releasing hormone) to more than 200 (in the case of follicle-stimulating hormone) amino acid residues and can have molecular weights as large as 30,000. All hormones secreted by the pituitary gland are peptide hormones, as are: Leptin from adipocytes Ghrelin from the stomach insulin from the pancreas 3. Steroid Steroid hormones are converted cholesterol. 187 from their parent compound, Vitamin D3 (steroid hormone) Mammalian steroid hormones can be grouped into five groups by the receptors to which they bind: 1. Gluco corticoids 2. Mineralo corticoids 3. Androgens 4. Estrogens 5. Progestagens Hormones as a signal Hormonal signaling across this hierarchy (chain of commands) involves the following: 1. Biosynthesis of a particular hormone in a particular tissue 2. Storage and secretion of the hormone 3. Transport of the hormone to the target cell(s) 4. Recognition of the hormone by an associated cell membrane or intracellular receptor protein. 5. Relay (send) and amplification of the received hormonal signal via a signal transduction process: This then leads to a cellular response. The reaction of the target cells may then be recognized 188 by the original hormone-producing cells, leading to a downregulation in hormone production. This is an example of a homeostatic negative feedback loop. 6. Degradation of the hormone. As can be inferred from the hierarchical diagram, hormone biosynthetic cells are typically of a specialized cell type, residing within a particular endocrine gland, such as thyroid gland, ovaries, and testes. Hormones exit their cell of origin via exocytosis or another means of membrane transport. The hierarchical model is an over simplification of the hormonal signaling process. Cellular recipients of a particular hormonal signal may be one of several cell types that reside within a number of different tissues, as is the case for insulin, which triggers a diverse (varied) range of systemic physiological affects. Different tissue types may also respond differently to the same hormonal signal. Because of this, hormonal signaling is elaborate (complicated) and hard to dissect (divided). Interactions with receptors Most hormones initiate a cellular response by initially combining with either a specific intracellular or cell membrane associated receptor protein. A cell may have several different receptors that recognize the same hormone and activate different signal transduction pathways, or 189 alternatively different hormones and their receptors may invoke (refer to) the same biochemical pathway. For many hormones, including most protein hormones, the receptor is membrane associated and embedded in the plasma membrane at the surface of the cell. The interaction of hormone and receptor typically triggers a cascade (flow) of secondary effects within the cytoplasm of the cell, often involving phosphorylation or de-phosphorylation of various other cytoplasmic proteins, changes in ion channel permeability, or increased concentrations of intracellular molecules that may act as secondary messengers (e.g. cyclic AMP). Some protein hormones also interact with intracellular receptors located in the cytoplasm or nucleus by an intracrine mechanism. For hormones such as steroid or thyroid hormones, their receptors are located intracellularly within the cytoplasm of their target cell. In order to bind their receptors these hormones must: Cross the cell membrane. Combined with receptor. The combined hormone-receptor complex moves across the nuclear membrane into the nucleus of the cell. Hormone binds to specific DNA sequences. Hormone is effectively amplifying or suppressing the action of certain genes. Hormones affecting protein synthesis. 190 However, it has been shown that not all steroid receptors are located intracellularly, some are plasma membrane associated. An important consideration, dictating the level at which cellular signal transduction pathways are activated in response to a hormonal signal is the effective concentration of hormone-receptor complexes that are formed. ENDOCRINE DISEASES A disease due to a disorder of the endocrine system is often called a "HORMONE IMBALANCE", but is technically known as an ENDOCRINOPATHY or ENDOCRINOSIS. Endocrine disease Classification and external resources Major endocrine glands. (Male left, female on the right.) 1. Pineal gland 2. Pituitary gland Thymus 5. Adrenal gland Ovary 8. Testes 191 3. Thyroid gland 6. Pancreas 4. 7. 7.METABOLISM METABOLISM is the set of chemical reactions that occur in living organisms in order to maintain life. These processes allow organisms to grow and reproduce, maintain their structures, and respond to their environments. Metabolism is usually divided into two categories. CATABOLISM breaks down large molecules, for example to harvest energy in cellular respiration. ANABOLISM , on the other hand, uses energy to construct components of cells such as proteins and nucleic acids. The chemical reactions of metabolism are organized into metabolic pathways, in which one chemical is transformed into another by a sequence of enzymes. Most of the structures that make up animals, plants and microbes are made from three basic classes of molecule: 1. amino acids 2. carbohydrates 3. lipids As these molecules are vital for life, metabolism focuses on making these molecules, in the construction of cells and tissues, or breaking them down and using them as a source of energy, in the digestion and use of food. 192 Many important biochemicals can be joined together to make polymers such as DNA and proteins. These macromolecules are essential parts of all living organisms. CATABOLISM Catabolism is the set of metabolic processes that break down large molecules. These include breaking down and oxidizing food molecules. The purpose of the catabolic reactions is to provide the energy and components needed by anabolic reactions. The exact nature of these catabolic reactions differ from organism to organism. The most common set of catabolic reactions in animals can be separated into three main stages: 1. Large organic molecules (proteins, carbohydrates or lipids) are digested into their smaller components outside cells. 2. These smaller molecules are taken up by cells and converted to smaller molecules, usually acetyl coenzyme A, which releases some energy. 3. The acetyl group of CoA is oxidized to H2O and CO2 in the citric acid cycle and electron transport chain, releasing the energy that is stored by reducing the coenzyme NAD+ (nicotinamide adenine dinucleotide) into NADH. 193 Digestion Macromolecules such as starch, cellulose or proteins cannot be rapidly taken up by cells and need to be broken into their smaller units before they can be used in cell metabolism. Several common classes of enzymes digest these polymers. These digestive enzymes include proteases that digest proteins into amino acids, as well as glycoside hydrolazes that digest polysaccharides into monosaccharides. Lipases digest lipids into fatty acids and glycerol. Animals secrete these enzymes from specialized cells in their guts. The amino acids or sugars released by these extracellular enzymes are then pumped into cells by specific active transport proteins. Energy from organic compounds Carbohydrate catabolism is the breakdown of carbohydrates into smaller units. Carbohydrates are usually taken into cells once they have been digested into monosaccharides. Once inside, the major route of breakdown is glycolysis, where sugars such as glucose and fructose are converted into pyruvate and some ATP is generated. 194 Pyruvate is an intermediate in several metabolic pathways, but the majority is converted to acetyl-CoA and fed into the citric acid cycle. Although some more ATP is generated in the citric acid cycle, the most important product is NADH, which is made from NAD+ as the acetyl-CoA is oxidized. An alternative route for glucose breakdown is the pentose phosphate pathway, which reduces the coenzyme NADPH and produces pentose sugars such as ribose, the sugar component of nucleic acids. Fats are catabolized by hydrolysis to free fatty acids and glycerol. The glycerol enters glycolysis and the fatty acids are broken down by beta oxidation to release acetyl-CoA, which then is fed into the citric acid cycle. Fatty acids release more energy upon oxidation than carbohydrates because carbohydrates contain more oxygen in their structures. Amino acids are either used to synthesize proteins and other biomolecules, or oxidized to urea and carbon dioxide as a source of energy. The oxidation pathway starts with the removal of the amino group by a transaminase. The amino group is fed into the urea cycle, leaving a deaminated carbon skeleton in the form of a keto acid. Several of these keto acids are intermediates in the citric acid cycle, for example 195 the deamination of glutamate forms α-ketoglutarate. The glucogenic amino acids can also be converted into glucose, through gluconeogenesis. ANABOLISM Anabolism is the set of constructive metabolic processes where the energy released by catabolism is used to synthesize complex molecules. In general, the complex molecules that make up cellular structures are constructed step-by-step from small and simple precursors. Anabolism involves three basic stages: 1. The production of precursors such as amino acids, monosaccharides, isoprenoids and nucleotides. 2. Their activation into reactive forms using energy from ATP. 3. The assembly of these precursors into complex molecules such as proteins, polysaccharides, lipids and nucleic acids. 196 Carbohydrates Metabolism In carbohydrate anabolism, simple organic acids can be converted into monosaccharides such as glucose and then used to assemble polysaccharides such as starch. The generation of glucose from compounds like pyruvate, lactate, glycerol, glycerate 3-phosphate and amino acids is called gluconeogenesis. Gluconeogenesis converts pyruvate to glucose-6-phosphate through a series of intermediates, many of which are shared with glycolysis. However, this pathway is not simply glycolysis run in reverse, as several steps are catalyzed by non-glycolytic enzymes. Polysaccharides and glycans are made by the sequential addition of monosaccharides by glycosyltransferase from a reactive sugar-phosphate donor such as uridine diphosphate glucose (UDP-glucose) to an acceptor hydroxyl group on the growing polysaccharide. 197 Glycolysis Digestion of Dietary Carbohydrates Dietary carbohydrate from which humans gain energy enter the body in complex forms, such as disaccharides and the polymers starch (amylose and amylopectin) and glycogen. The polymer cellulose is also consumed but not digested. The first step in the metabolism of digestible carbohydrate is the conversion of the higher polymers to simpler, soluble forms that can be transported across the intestinal wall and delivered to the tissues. Oxidation of glucose is known as glycolysis. Glucose is oxidized to either lactate or pyruvate. Under aerobic conditions, the dominant product in most tissues is pyruvate and the pathway is known as aerobic glycolysis. When oxygen is depleted, as for instance during prolonged vigorous exercise, the dominant glycolytic product in many tissues is lactate and the process is known as anaerobic glycolysis. The Energy Derived from Glucose Oxidation 198 Glucose + 2 ADP + 2 NAD+ + 2 Pi 2 Pyruvate + 2 ATP + 2 NADH + 2 H+ The NADH generated during glycolysis is used to fuel mitochondrial ATP synthesis via oxidative phosphorylation, producing either two or three equivalents of ATP. The net yield from the oxidation of 1 mole of glucose to 2 moles of pyruvate is, therefore, either 6 or 8 moles of ATP. Complete oxidation of the 2 moles of pyruvate, through the Citric Acid Cycle (TCA cycle) yields an additional 30 moles of ATP; the total yield, therefore being either 36 or 38 moles of ATP from the complete oxidation of 1 mole of glucose to CO2 and H2O. The citric acid cycle is part of a metabolic pathway involved in the chemical conversion of carbohydrates, fats and proteins into carbon dioxide and water to generate a form of usable energy. 199 The Individual Reactions of Glycolysis The pathway of glycolysis can be seen as consisting of 2 separate phases. The first is the chemical priming phase requiring energy in the form of ATP. The second is considered the energy-yielding phase. In the first phase, 2 equivalents of ATP are used to convert glucose to fructose 1,6-bisphosphate (F1,6BP). In the second phase, fructose 1,6-bisphosphate (F1,6BP) is degraded to pyruvate, with the production of 4 equivalents of ATP and 2 equivalents of NADH. 200 201 Pathway of Glycolysis from glucose to pyruvate (Lactate). Embden-Mayerhof- دورة امدن مايرهوف 202 Anaerobic Glycolysis Under aerobic conditions, pyruvate in most cells is further metabolized via the TCA cycle. Under anaerobic conditions and in erythrocytes under aerobic conditions, pyruvate is converted to lactate by the enzyme lactate dehydrogenase (LDH), and the lactate is transported out of the cell into the circulation. The conversion of pyruvate to lactate, under anaerobic conditions, provides the cell with a mechanism for the oxidation of NADH (produced during the G3PDH reaction) to NAD+; which occurs during the LDH catalyzed reaction. Aerobic glycolysis generates substantially more ATP per mole of glucose oxidized than does anaerobic glycolysis. The utility of anaerobic glycolysis, to a muscle cell when it needs large amounts of energy, stems from the fact that the rate of ATP production from glycolysis is approximately 100X faster than from oxidative phosphorylation. 203 Gluconeogenesis Gluconeogenesis is the biosynthesis of new glucose, (i.e. not glucose from glycogen). The production of glucose from other metabolites is necessary for use as a fuel source by the brain, testes, erythrocytes and kidney medulla since glucose is the sole energy source for these organs. During starvation, however, the brain can derive energy from ketone bodies which are converted to acetyl-CoA. 204 205 206 Substrates for Gluconeogenesis Lactate: Lactate is a predominate source of carbon atoms for glucose synthesis by gluconeogenesis. During anaerobic glycolysis in skeletal muscle, pyruvate is reduced to lactate by lactate dehydrogenase (LDH). LDH Pyruvate Lactate This reaction serves two critical functions during anaerobic glycolysis: 1. LDH reaction requires NADH and yields NAD+ which is then available for use by the glyceraldehyde-3- phosphate dehydrogenase reaction of glycolysis. 2. The lactate produced by the LDH reaction is released to the blood stream and transported to the liver where it is converted to glucose. The glucose is then returned to the blood for use by muscle as an energy source and to replenish glycogen stores. This cycle is termed the Cori cycle. 207 Pyruvate: Pyruvate, generated in muscle and other peripheral tissues, can be transaminated to alanine which is returned to the liver for gluconeogenesis. The transamination reaction requires an α-amino acid as donor of the amino group, generating an α-keto acid in the process. This pathway is termed the cycle. 208 glucose-alanine The glucose-alanine mechanism for cycle muscle is, to therefore, eliminate an nitrogen indirect while replenishing يزززودits energy supply. However, the major function of the glucose-alanine cycle is to allow non-hepatic tissues to deliver the amino portion of catabolized amino acids to the liver for excretion as urea. Within the liver the alanine is converted back to pyruvate and used as a gluconeogenic substrate (if that is the hepatic requirement) or oxidized in the TCA cycle. The amino nitrogen is converted to urea in the urea cycle and excreted by the kidneys. 209 Citric Acid Cycle The citric acid cycle — also known as the tricarboxylic acid cycle (TCA cycle), the Krebs cycle, is a series of enzymecatalysed chemical reactions of central importance in all living cells that use oxygen as part of cellular respiration. In eukaryotes, the citric acid cycle occurs in the matrix of the mitochondrion. In aerobic organisms, the citric acid cycle is part of a metabolic pathway involved in the chemical conversion of carbohydrates, fats and proteins into carbon dioxide and water to generate a form of usable energy. Other relevant reactions in the pathway include those in glycolysis and pyruvate oxidation before the citric acid cycle, and oxidative phosphorylation after it. In addition, it provides precursors for many compounds including some amino acids and is therefore functional even in cells performing fermentation. The TCA cycle showing enzymes, substrates and products. The GTP generated during the succinate thiokinase (succinyl-CoA synthetase) reaction is equivalent to a mole of ATP by virtue of the presence of nucleoside diphosphokinase. The 3 moles of NADH and 1 mole of FADH2 generated during each round of the cycle feed into the oxidative phosphorylation pathway. Each mole of NADH leads to 3 moles of ATP and each mole of FADH2 leads to 2 moles of 210 ATP. Therefore, for each mole of pyruvate which enters the TCA cycle, 12 moles of ATP can be generated. IDH = isocitrate dehydrogenase. α-KGDH = α-ketoglutarate dehydrogenase. MDH = malate dehydrogenase. Place mouse over cycle intermediate names to see their structures. 211 CITRIC ACID CYCLE (TCA CYCLE) Krebs دورة 212