Diffusion Confusion Mark Anderson Fran Butek Andy
advertisement

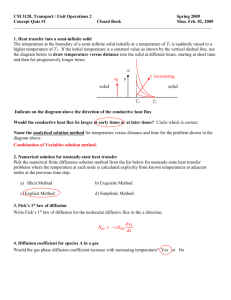
Diffusion Confusion Mark Anderson Fran Butek Andy Dettinger Dan Hecker Mark Osegard Overview •Diffusion •A Biological Ohm’s Law •Facilitated Diffusion •Muscle Respiration Mechanisms of Cellular Homeostasis ho·me·o·sta·sis: The ability or tendency of an organism or cell to maintain internal equilibrium by adjusting its physiological processes. Cellular Homeostasis Cell Membrane The Cell Membrane Acts as a boundary that separates the internal working of the cell from its external environment. It allows free passage of some material and restricts the passage other. In this way the cell membrane regulates the passage of materials into and out of the cell. The Cell Membrane (cont.) It consists of a double layer of phospholipid molecules. Examples of phospholipids include lecithin, cephalins, phosphoinositides (in brain), and cardiolipin (in heart). The membrane contains water-filled pores and protein-lined pores, called channels. These channels allow passage of specific molecules. The Cell Membrane (cont.) The membrane’s intracellular and extracellular environment are made up primarily of NaCl and KCl The cell membrane works as a barrier to the free flow of these ions. The membrane also maintains concentration differences between ions, and acts as a barrier to water flow. Cellular Transportation Two Types of Transportation Molecules can be transported across the cell membrane by passive or active processes. Active process requires the expenditure of energy. Cellular Transportation (cont.) The passive process results from the, random movement of the molecules. There are three passive transport mechanisms 1) Osmosis - is the process by which water is transported through the cell membrane 2) Simple diffusion - accounts for the passage of small molecules through pores and of lipid-soluble molecules through the bilipid layer. 3) Carrier-mediated diffusion - occurs when a molecule is bound to a carrier molecule that moves readily through the membrane. Making the Connection The cell membrane is a medium where diffusion takes place. Our study this semester of diffusion, and what will be shown in the following slides, is the diffusion where the membrane is a group of cells, such as muscle tissue. Hammering out the Diffusion Equation Compartmental Analysis Diffusion through a Region Divergence Theorem The Diffusion Equation Fick’s Law Refined Diffusion Equation Compartmental Analysis X in X X out X is some variable of interest A change in X = X in – X out in derivative terms: dx = xin (t ) − xout (t ) dt What is Diffusion? Definition – the process by which a substance disperses within an ambient medium over time Modeled using compartmental analysis Net change of substance at a point = (inflow rate of substance) – (outflow rate of the substance) Diffusion through a Region The region may contain sources and sinks. ( x, y , z ) ∈ Ω Modeling Diffusion (the nuts and bolts) u = u(x, y, z, t) = density at a point in the region f = f(x, y, z, t) = production at a point (a density) in the region due to sources and sinks J = J(x, y, z, t) = flux density at a point Some Calculus (u )dV = total mass in region Ω d dt ( u ) dV = rate of mass change in region Ω ( f ) dV = total production of mass in Ω region due to sources and sinks The Confusing Quantity of Flux J = J(x, y, z, t) = flux density at a point (density x velocity) The units of flux are: mass area * time small flux in the negative x direction large flux in the positive x direction What can we do with Flux? Mass leaving the region through the boundary will be the component of the flux that is in the direction of the surface normal J ⋅ n = total flux out of the region in the direction of at a point on the n boundary ( J ⋅ n )dA ∂Ω = total flux (accumulation of mass) out of the region through the boundary J ( x, y , z , t ) n Putting Together the Pieces Using compartmental analysis we can write an equation relating all the components we have developed thus far. rate of substance change = amount produced in the region – amount that escapes the region d (u )dV = ( f )dV − ( J ⋅ n )dA dt Ω Ω ∂Ω Divergence Theorem (Gauss’s Theorem) a Method of modifying the flux integral gradient ∂ ∂ ∂ ∇=( , , ) ∂x ∂y ∂z The amount of mass that overflows the boundary must be equal to the total change in mass within the boundary. The Bathtub Overflow Theorem ( J ⋅ n)dA = (∇ ⋅ J )dV ∂Ω Ω Johann Carl Friedrich Gauss Born: 30 April 1777 Brunswick, Germany Died: 23 Feb 1855 Göttingen, Germany Simplification d (u ) dV = ( f ) dV − ( J ⋅ n ) dA dt Ω Ω ∂Ω Divergence Theorem ∂u ( ) dV = ( f ) dV − (∇ ⋅ J ) dV ∂t Ω Ω Ω ∂u ( ) dV = ( f − ∇ ⋅ J ) dV ∂t Ω Ω Almost there… ∂u ( ) dV − ( f − ∇ ⋅ J ) dV = 0 ∂t Ω Ω ∂u ( − f + ∇ ⋅ J ) dV = 0 ∂t Ω If you integrate over all subregions of “OMEGA” and get zero, the expression integrated must also be equal to zero ∂u − f +∇⋅J = 0 ∂t The Unrefined Diffusion Equation ∂u = f − ∇⋅ J ∂t density change at a point over time = density production at that point per unit time – the divergence (the rate of flow from that point) Gradient Fick’s Law ∂ ∂ ∂ ∇=( , , ) ∂x ∂y ∂z During diffusion we assume particles move in the direction of least density. They move down the concentration gradient In mathematical terms we will assume J = − D∇u Where D is a constant of proportionality called the Diffusion Coefficient Adolph Fick Born: 1829 Cassel, Germany Died: 1879 Ockham’s Razor ∂u − f +∇⋅J = 0 ∂t When faced with a choice between two things, choose the simpler. If we assume D is constant and substitute the new expression we found for flux using Fick’s Law we can remove the flux density component of our diffusion equation as follows: ∂u = f − ∇⋅ (−D∇u) ∂t ∂u = f + D∇⋅ ∇u ∂t ∂u = f + D∇2u ∂t William of Ockham Born: 1288 Ockham, England Died: 9 April 1348 Munich, Bavaria The Laplacian ∂ ∂ ∂ ∇ =( 2 + 2 + 2) ∂x ∂y ∂z 2 2 2 2 Pierre-Simon Laplace Born: 23 March 1749 Normandy, France Died: 5 March 1827 Paris, France The Refined Diffusion Equation ∂u 2 = f + D∇ u ∂t A Biological Ohm’s Law (a formulation of the relationship of voltage, current, and resistance) Ohm’s law is defined most simply using three terms V, I, and R. V=IxR Where: V = voltage measured in volts I = current measure in amperes R = resistance measure in Ohms So what does Ohm’s law have to do with Diffusion? Using Fick’s Law we can derive a chemical counterpart of Ohm’s Law that can be used to describe diffusion through a membrane of thickness L. large chemical reservoir cl membrane L large chemical reservoir cr As you can see from the model, this membrane has a concentration of cl on the left and a concentration of cr on the right Applying the diffusion Equation Let c(x,t) denote the concentration at x at time t in previous one dimensional model. If we assume no local production we can model the concentration using the diffusion equation and write: 2 ∂c ∂c =D 2 ∂t ∂x Boundary conditions c(0,t) = cl c(L,t) = cr Simplification by Assumption In order for us to simplify our lives and the following equation, we ignore time and assume it to be at steady-state, therefore the change in concentration will fall off to zero: ∂c =0 ∂t Using the steady state equation and our equation from the previous slide we can write: D 2 ∂ c ∂x 2 =0 One dimensional Laplacian A Linear Model Dividing by D and integrating with respect to x we get ∂c =a ∂x where “a” is a constant. Then we integrate with respect to x one more time and see that c(x) is linear in steady state: c ( x ) = ax + b Boundary Conditions c(0) = cl and c( L) = cr b = cl cr = aL + b we define a and b to be: cr − cl a= L So recalling the linear formula we found previously c ( x ) = ax + b we plug in the a and b values defined above. The resulting equation then becomes: x c( x) = cl + (cr − cl ) L Differentiating ∂c cr − cl = ∂x L differentiating with respect to x Now recalling Fick’s Law, with a one dimensional gradient (aka a derivative), which Mark Anderson used earlier we write: Note that in one dimension J is scalar, rather than a vector as before Plugging in: ∂c ∂x J = − D∇c ∂c = −D ∂x cl − cr J =D L Made It! We can rewrite as D J = (cl − cr ) L Using the equation from my first slide, solved for I We can see a correspondence between our 1 I= V R model for flux and Ohm’s Law. The ratio L / D is similar to “resistance” of the membrane and subsequently the ratio D / L is the conductance, or the permeability of the membrane. J = I(current) D/L = 1/R(conductivity) (cl-cr) = V(voltage drop) A Biological Ohm’s Law D J = (cl − cr ) L Ohm’s Law is the simplest diffusion scenario. It is conducted in one dimension when there are no reactions occurring. Reaction-Diffusion Systems Reactants in an enzymatic reaction are free to diffuse, so that one must keep track of the effects of both diffusion and reaction. An example in which both diffusion and reaction play a role is known as facilitated diffusion. Facilitated Diffusion Facilitated diffusion occurs when the flux of a chemical is increased by a reaction that takes place in the diffusing medium. An example of facilitated diffusion occurs with the flux of oxygen in muscle fibers. Oxygen in Muscle Fibers In muscle fibers, oxygen is bound to myoglobin, a much larger molecule, and is transported as oxymyoglobin. This transport is greater than that of free oxygen, and hence the flux of oxygen is enhanced. Whoa! I know what you are thinking: “ain’t that kind of weird since myoglobin has a much larger molecular weight (16,890amu) than oxygen (32amu), and therefore a much smaller diffusion coefficient!?” Doxygen=4.4*10-7cm2/s Dmyglobin=1.2*10-5cm2/s A Simple Model To help us understand this phenomenon, consider a slab reactor (or reservoir) in which oxygen and myoglobin react to form oxymyoglobin. Myoglobin is not allowed to leave the slab in a free or bound form, but oxygen may flow in and out freely. Slab Reactor k+ → ← k O2 + Mb large chemical reservoir of pure O2 x=0 slab 0<x<L − MbO2 large chemical reservoir of pure O2 x=L Definitions Let s = s(x,t) denote the concentration of oxygen at a point x and time t Let e=e(x,t) denote the concentration of myoglobin at a point x and time t Let c=c(x,t) denote the concentration of oxymyoglobin at a point x and time t Applying the Diffusion Equation If we let f denote the rate of uptake of oxygen into oxymyoglobin, we can use the diffusion equation to model the concentrations s = [O2], e = [Mb], c = [MbO2] as shown below: Note that f will play the role of either a source or a sink depending its sign. ∂ s ∂s − f = Ds 2 ∂t ∂x 2 ∂ e ∂e − f = De 2 ∂x ∂t 2 ∂ c ∂ c 3 instances of + f = Dc the diffusion 2 ∂t ∂x equation 2 Assumptions Myoglobin and oxymyoglobin are nearly identical in molecular weight and structure, therefore we assume: De ≈ Dc We will assume that as we approach the boundaries of the slab the changes in myglobin and oxymyoglobin taper off, giving us the boundary conditions at x=0 and x=L to be : ∂e ∂c = =0 ∂x ∂x Compartmental Analysis O2 Mb k+ se MbO2 k-c k+ → ← k O2 + Mb − MbO2 Diagram Equation From the compartmental diagram we can write the following equation dc = k + se − k − c dt This equation describes the uptake of oxygen into oxymyoglobin, hence we can write: f = k + se − k − c Elimination Because total enzyme in the slab reactor is conserved we know that in the total free enzyme + the total bound enzyme must = the total enzyme before reactions began at time t=0 e + c = e0 e = e0 − c Since e actually depends on c we can eliminate the following equation from our system ∂e ∂ e = De 2 − f ∂t ∂x 2 Quasi Steady State Recall our current system of equations, now that e has been eliminated as a variable ∂s ∂2s = Ds − f 2 ∂t ∂x ∂c ∂ 2c + f = Dc 2 ∂t ∂x f = k + se − k − c At Quasi Steady state we assume rates of formation and rates of breakdown are the same, and that at steady state the rates of change are 0. ∂s ∂c + =0 ∂t ∂t Substituting Taking our previous equation and substituting from our current system of equations we get 0 = Ds ∂2s ∂x 2 + Dc ∂ 2c ∂x 2 ∂c ∂ ∂s + Dc Ds 0= ∂x ∂x ∂x ∂s ∂c + Dc =c Ds ∂x ∂x Fick’s Law Revisited Recall that by Fick’s law Flux = -(diffusion coefficient)*(spatial derivative of density) Using this we can model the total oxygen flux J=J(x) at a point as ∂c ∂s + Dc J = − Ds ∂x ∂x Integrating ℑ= L 0 ds dc − Ds + Dc dx dx dx ℑ = − Ds (s0 − s L ) − Dc (c0 − cL ) The quantities s0 and sl are known , but cl and c0 are not! We can find cl and c0 these by using nondimensionalization . Nondimensionalization To further simplify and understand this system of equations we need to define some new quantities. k+ σ = s k− c u= e0 x = Ly Scaled Substrate Scaled Complex Rescaled spatial variable More Nondimensionalization If we use these with our current system of equations we arrive at the following equation: ε1σ yy = σ (1 − u ) − u = −ε 2u yy where ε1 = Ds 2 e0 k + L ε2 = Dc 2 k− L Using Experimental Data Experimental data suggests the following epsilon values ε1 = 1.5 *10 −7 ε 2 = 8.2 *10 These number are small enough that we can approximate them with zero, leaving us with the equation to solve for u σ (1 − u ) − u = 0 σ u= σ +1 −5 Back Substituting By back substituting and going through a LOT of algebra we can get the following k+ s k− σ u= = σ +1 k+ k− s +1 c = e0 c s = e0 s + k − k+ cL and c0 Revealed! k− Let K := k+ e0 s c= s+K Now, if we use our boundary conditions of c(0)=c0 and c(L)= cl we get e0 s 0 c0 = s0 + K e0 s L cL = sL + K Total Oxygen Flux Dc Ds (s0 − sL ) + (c0 − cL ) ℑ= L L s0 Dc Ds sL ) ℑ = (s0 − sL ) + e0 ( − L K + s0 K + sL L = Back substituting are new values for cL and c0 into our equation for the total flux of oxygen in the slab leads to the following D e0 K Ds (s0 − sL )(1+ c ) Ds (s0 + K)(sL + K) L Ds = (1 + µρ )( s 0 − s L ) L D c e0 ρ = Ds K K2 µ= ( s0 + K )(sL + K ) What does it all mean!? Dan H. introduced us to the concept of cellular homeostasis. Mark A. developed the Diffusion Equation. Mark O. developed a Biological version Ohm’s Law. Andy D. developed a general model of Facilitated Diffusion, and walked us through a case study of a Reaction Diffusion system. Muscle Respiration Definition – Muscle respiration refers to the ability of muscle to extract oxygen from the blood being delivered to it by the heart and circulatory system. This can be modeled using the Reaction Diffusion system. A Cylindrical Model z Cylindrical Coordinates: (r, , z) r y x Muscle Fiber: a cylindrical model Assumption and Analogy Physical Assumptions: i) The Oxygen concentration is fixed on the outside of the “fiber”. ii) At steady state the oxygen (s) and oxymyoglobin (c) are distributed radially and symmetrically about the z axis. In effect, the substrate only depends on r, rather than (r, , z). Recall the Diffusion Equation ∂u 2 = D∇ u + f ∂t Where u is the concentration at a point and time. u = u ( x, y , z , t ) and the Laplacian of u is: ∂ u ∂ u ∂ u ∇ u= 2 + 2 + 2 ∂x ∂y ∂z 2 2 2 2 Laplacian in Cylindrical Coordinates ∂ 1 ∂ 1 ∂ ∂ + 2 ∇ = + r 2 2 r ∂r ∂r r ∂θ ∂z 2 2 2 Using our assumption of radial symmetry the Laplacian can be rewritten as: ∂ 1 ∂ ∇ = r r ∂r ∂r 2 In the Absence of Oxygen Consumption… ∂s 2 = Ds ∇ s − f ∂t ∂c 2 = Dc ∇ c + f ∂t The rate of change of oxygen concentration with respect to time = diffusion coefficient of the oxygen * Laplacian of the oxygen concentration – uptake of oxygen into oxymyoglobin. Note: f denotes the rate of uptake of oxygen into oxymyoglobin. Oxygen Consumption in Steady - State ∂s 2 2 = Ds ∇ s − f 0 = Ds ∇ s − f ∂t 1 ∂ ∂s Expanding the 0 = Ds r −f Laplacian r ∂r ∂r Accounting for oxygen depletion 1 ∂ ∂s r − f −g 0 = Ds r ∂r ∂r Oxymyoglobin in Steady – State ∂c =0 ∂t Expanding the Laplacian 0 = Dc ∇ c + f 2 1 ∂ ∂c +f r 0 = Dc r ∂r ∂r Modeling Muscle Respiration Describing the rate of change of oxygen concentration during diffusion: 1 ∂ ∂s r − f −g 0 = Ds r ∂r ∂r Describing the rate of change of oxymyoglobin concentration during diffusion: 1 ∂ ∂c 0 = Dc r +f r ∂r ∂r Boundary Conditions s = sa ∂c =0 ∂r At r = a where a is the maximum radius at r = a ∂s ∂c = =0 ∂r ∂r at r = 0 -a 0 a What’s our Conclusion? We’ve used mathematical models, in particular the diffusion equation, to explain the metabolic process of oxygen consumption in muscle respiration. The final model we developed can be used in combination with asymptoptic methods to discern information about the steady state behavior such processes. We will be continuing our study of this topic next semester. Special Thanks Dr. Steve Deckelman - (patience and pizza) Dr. Ann Parsons - (expertise) Mr. Adolph Fick - (great flux model) Sir William of Ockham - (razor) Mr. Johann Carl Friedrich Gauss - (divergence) Mr. Pierre-Simon Laplace - (Laplacian) References Mathematical Physiology Keener and Sneyd Springer-Verlag, 1998 Thomas’ CALCULUS Revised by Finney, Weir and Giordano Addison Wesley Longman, 2001