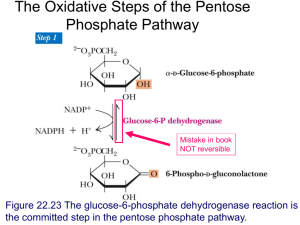
18_gluconeo-and
advertisement

GLYCOGEN METABOLISM Glycogen: a highly branched polymer of glucose. Chains have glycosidic links α 14. Branches are linked α 16. Glucose stored in polymeric form as glycogen mostly in the liver and skeletal muscle. Glucose can be rapidly delivered to blood stream when needed upon degradation of glycogen. = glycogenolysis Enough glucose and energy triggers synthesis of glycogen. = glycogenesis Glycogenesis = GLYCOGEN BIOSYNTHESIS ============================== Glucose is transported into the cell by a specific glucose transporter and immediately phosphorylated. ============================== -- Most of the glucose in a cell is in the form of glucose-6-phosphate. 1- Conversion of glucose-6-phosphate to glucose-1-phosphate Enzyme = phosphoglucomutase α-D-glucose-6α-D- glucose-1phosphate phosphate reversible reaction allows G1P conversion to G6P in glycogenolysis mechanism involves phosphorylated enzyme intermediate and glucose-1,6 bisphosphate bound intermediate similar to phosphoglycerate mutase 2- Synthesis of Uridine Diphosphoglucose Enzyme = UDP-glucose pyrophosphorylase Reaction: glucose-1-phosphate + UTP UDP-glucose + PPi Then PPi 2 Pi the commiting step Phosphoryl transfer -- UTP is the energy equivalent of ATP -- energy is used to activate glucose -- two phosphates from UTP are lost as PPi PPi is broken down PPi 2 Pi driving the reaction to the right 3- Glycogen synthesis Enzyme = glycogen synthase UDP-glucose + (glucose)n UDP+(glucose)n+1 glycogen 4- Branching Enzyme = branching enzyme introduces branching by transferring a teminal fragment of 6-7 residues from a growing chain to a 6-position farther back in the chain. makes a branch with an α (16) link and two ends to add glucose. branching accelerates the rate of glucose release during degradation. new 1,6 branching bond enzyme -- GLYCOGENOLYSIS-DEGRADATION OF GLYCOGEN 1. Release of glucose-1-phosphate Enzyme = glycogen phosphorylase nonreducing + Pi ends glucose-1- + phosphate always acts at nonreducing end 1,4 glycosidic link is cleaved by phosphorylysis with retention of energy potential in the phosphate ester of glucose-1-phosphate. stops at fourth glucose from a 1,6 branch point contrast with enzymes acting on starch and glycogen in the gut, which yield sugars, not sugar phosphates, as products. activated by phosphorylation, regulated by glucagon and epinephrine 2. Debranching - two parts Enzyme = debranching enzyme (both) (16) link transferase Transfers chain of three glucoses to any nonreducing end (16) link debranching enzyme (glucosidase) + = glucose 1,6 linkage cleaved glycogen phosphorylase or phosphorylase for short glucose-1-phosphate -- phosphoglucomutase then yields glucose-6-phosphate, which can enter glycolysis. Metabolic Regulation of Mammalian Glycogen Levels -- Glycogen reserves are the most immediately available large source of metabolic energy for mammals. -- Storage and utilization are under dietary and hormonal control. Primary hormones = -- epinephrine (adrenaline) = “fight-or-flight” -- glucagon -- insulin Primary enzyme targets in glycogen metabolism= glycogen phosphorylase and glycogen synthase. The actions of the hormones are indirect. ======== HORMONES =========== Glucagon - low glucose levels -- Acts primarily in liver. -- A polypeptide hormone produced in α-cells of the islets of Langerhan of the pancreas. -- Receptors on surface of liver cells. -- Stimulates glycogen breakdown & inhibits glycogenesis. -- Glucagon also blocks glycolysis & stimulates gluconeogenesis. Epinephrine - low glucose levels -- Acts primarily on skeletal muscle. -- Receptors on surface of cells. -- Stimulates glycogen breakdown & inhibits glycogenesis. Glucagon and epinephrine both stimulate intracellular pathway via increasing levels of cAMP. Insulin -- High levels of glucose induce release of insulin from β-cells of islets of Langerhan in the pancreas. -- Insulin is polypeptide hormone. -- Detected by receptors at surface of muscle cells. -- Increases glycogenesis in muscle. cAMP Cascade -- A cyclic AMP cascade is used by both epinephrine and glucagon. -- A cascade is a mechanism in which enzymes activate other enzymes sequentially usually leading to an amplification of an initial signal. Epinephrine/Glucagon Cascade Regulating Glycogen Metabolism Glycogen Storage Diseases: -- A family of serious, although not necessarily fatal, diseases caused by mutations in the enzymes involving in glycogen storage and breakdown. -- GLUCONEOGENESIS -- Definition: the biosynthesis of glucose from simpler molecules, primarily pyruvate and its precursors. pyruvate lactate some amino acid skeletons TCA cycle intermediates The gluconeogenesis pathway is similar to the reverse of glycolysis but differs at critical sites. control of these opposing pathways is reciprocal so that physiological conditions favoring one disfavor the other and vice versa. General principles of metabolic control -- a) pathways are not simple reversals of each other and b) under reciprocal control Why do we produce glucose? a) need to maintain glucose levels in a narrow range in blood. b) some tissue-- brain, erythrocytes, and muscles in exertion use glucose at a rapid rate and sometimes require glucose in addition to dietary glucose. [The brain and erythrocytes can use only glucose as a source of energy.] Where is glucose synthesized? The liver comes to rescue. The liver is the major location for gluconeogenesis. Sources of precursors: lactate from muscle, amino acids from diet or breakdown of muscle protein during starvation, propionate from breakdown of fatty acids and amino acids and glycerol from certain fats. In glycolysis, there are three irreversible kinase reactions at control points involving: hexokinase, phosphofructokinase, and pyruvate kinase Cost: The production of glucose is energy expensive. Input: 2 pyruvate + 4 ATP + 2 GTP + 2 NADH Output: glucose + 4 ADP + 2 GDP + 2 NAD++ 6 Pi Gluconeogenesis - beginning GDP + CO2 GTP Phosphoenolpyruvate phosphoenolpyruvate carboxykinase BYPASS 1 Oxaloacetate Some amino acids ADP + Pi ATP + CO2 pyruvate carboxylase (pyruvate kinase) PYRUVATE (3C) lactate, alanine, other amino acids fructose 1,6-bisphosphate aldolase triose phosphate isomerase dihydroxyacetone phosphate glyceraldehyde 3-phosphate dehydrogenase glyceraldehyde 3-phosphate NAD+ +Pi NADH +H+ 1,3-bisphosphoglycerate ADP phosphoglycerate kinase ATP 3-phosphoglycerate phosphoglyceromutase enolase 2-phosphoglycerate phosphoenolpyruvate GLUCOSE glucose Pi BYPASS 3 6-phosphatase (hexokinase) Glucose 6-phosphate phosphoglucoisomerase Fructose 6-phosphate BYPASS 2 fructose 1,6 bisphosphatase Pi (phosphofructokinase) Fructose 1,6-bisphosphate Bypass number 1. Pyruvate to phosphoenolpyruvate. This bypasses pyruvate kinase. Complex scheme. a) pyruvate to oxaloacetate Enzyme = pyruvate carboxylase located inside mitochondria. Only this enzyme of the gluconeogenesis pathway is mitochondrial. Reaction: pyruvate + CO2 + ATP + H2O oxaloacetate + ADP + Pi -- Carboxylations involving CO2 almost always use the vitamin biotin as a cofactor. -- Subreactions: Enz-biotin + ATP + CO2 + H2O Enz-carboxybiotin + ADP + Pi Enz-carboxybiotin + pyruvate Enz-biotin + oxaloacetate -- pyruvate carboxylase - acetyl-CoA is positive modulator absolutely required for activity higher acetyl-CoA indicates that adequate carbon levels available for TCA cycle to provide energy glucose can be synthesized and exported from liver. oxaloacetate important in the citric acid cycle, which is more mitochondrial. For gluconeogenesis, oxaloacetate must leave the mitochondria because all the rest of the gluconeogenesis enzymes are in the cytosol. mitochondrial membranes are nearly impermeable to oxaloacetate. So how does it get out? c) The GTP-dependent decarboxylation of oxaloacetate Enzyme = PEP carboxykinase Cytosolic enzyme, as all others oxaloacetate + GTP PEP + CO2 + GDP uses GTP, not ATP. CO2 added is lost in this step. NET: pyruvate + ATP + GTP PEP + ADP + GDP + Pi High cost = two energy rich phosphates so a total of four high energy bonds are already utilized here per glucose to be synthesized. Then uses glycolytic enzymes in steps back to fructose-1,6-bisphosphate Bypass number 2. Fructose-1,6bisphosphate to fructose-6-phosphate Enzyme = fructose-1,6-bisphosphatase Reaction: fructose-1,6-bisphosphate + H2O fructose-6-P + Pi bypasses phosphofructokinase a simple hydrolysis. highly exergonic, irreversible enzyme is highly regulated F6P isomerizes to glucose-6-phosphate via phosphoglucoisomerase Bypass number 3. Glucose-6-phosphate to glucose. Enzyme = glucose-6-phosphatase Reaction: glucose-6-phosphate + H2O glucose + Pi bypasses hexokinase highly exergonic, irreversible not present in muscle *************************************** Total Energy Cost = 6 high energy bonds used per glucose synthesized. four more than produced in glycolysis. These four are needed to convert pyruvate to PEP. ************************************** CONTROL: -- gluconeogenesis serves as an alternative source of glucose when supplies are low and is largely controlled by diet. -- high carbohydrate in meal reduce gluconeogenesis and starvation increases. -- key enzymes targeted. -- gluconeogenesis and glycolysis are controlled in reciprocal fashion. REGULATION OF GLUCONEOGENESIS: Key enzymes: 1. PYRUVATE CARBOXYLASE. activated by acetyl-CoA (required) vs. pyruvate kinase, inhibited by acetyl CoA. high levels of acetyl-CoA signals that enough carbon substrate available for citric acid cycle. pyruvate kinase is also inhibited by ATP and the liver form by alanine. 2. FRUCTOSE 1,6 BISPHOSPHATASE Strongly inhibited by AMP Strongly inhibited by fructose- 2,6-bisphosphate Recall that the reciprocal enzyme, phosphofructokinase, in glycolysis is strongly activated by AMP and fructose-2,6-bisphosphate. Fructose-2,6-bisphosphate is the most important regulator of glycolysis and gluconeogenesis through its reciprocal effects on fructose 1,6-bisphosphatase and phosphofructokinase. • F-2,6-BP synthesis is controlled by cAMP •F-2,6-BP is formed from F-6-P by PFK-2 3. PHOSPHOFRUCTOKINASE-2/ FRUCTOSE BISPHOSPHATASE-2 Bifunctional enzyme fructose-6-phosphate + ATP fructose-2,6-bisphosphate fructose-2,6-bisphosphate fructose-6-phosphate + Pi Enzyme highly regulated to control levels of F 2,6-P • In the heart muscle cell, cAMP activate • In skeletal muscle cell has no cAMP reg 3. PHOSPHOFRUCTOKINASE-2/ FRUCTOSE BISPHOSPHATASE-2 Bifunctional enzyme • In the liver, cAMP inhibits PFK-2, activating FBPase-2 • In the heart muscle cell, cAMP activates rather than inhibits PFK-2, since gluconeogenesis is not possible. • In skeletal muscle cell has no cAMP regulation site. Glucagon: hormone released when glucose levels are low. -- elevates blood glucose levels -- increases intracellular levels of cAMP in liver and elsewhere -- cAMP activates protein kinase A = cAMP-dependent protein kinase -- stimulating gluconeogenesis and glycogenolysis Summary of Gluconeogenesis purpose- alternative source of glucose rather than dietary carbohydrates or glycogen breakdown primary precursers are lactate, pyruvate, glycerol, part of fatty acids and certain amino acids (glucogenic) 3 essentially irreversible steps of glycolysis are bypassed regulated via pyruvate carboxylase, fructose 1,6 bisphosphatase, and phosphofructokinase-2/fructose bisphosphatase-2 glucose cannot be made from acetyl CoA