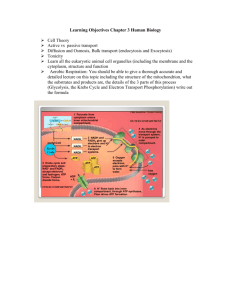
The reactions of the citric acid cycle.

KAPITOLA 9
Energetický metabolismus II
• cyklus trikarboxylových kyselin
• dýchací řetězec
• aerobní fosforylace
Catabolism of proteins, fats, and carbohydrates occurs in the three stages of cellular respiration.
Stage 1: Oxidation of fatty acids, glucose, and some amino acids yields acetyl-CoA.
Stage 2: Oxidation of acetyl groups via the citric acid cycle includes four steps in which electrons are abstracted.
Stage 3: Electrons carried by NADH and FADH
2 are funneled into a chain of mitochondrial (or plasma membrane-bound, in bacteria) electron carriers the respiratory chain - ultimately reducing O
2 to
H
2
O. This electron flow drives the synthesis of
ATP, in the process of oxidative phosphorylation.
Each turn of the cytric acid cycle produces three NADH and one
FADH
2 as well as one GTP (or
ATP ).
Two CO
2 are released in oxidative decarboxylation reactions.
The reactions of the citric acid cycle.
The carbon atoms are those originally derived from the acetate of acetyl-CoA in the first turn of the cycle. These carbons are not the ones released as CO
2 in the first turn. Note that in fumarate, the two-carbon group derived from acetate can no longer be specifically denoted; because succinate and fumarate are symmetric molecules, C-1 and C-2 are indistinguishable from C-4 and C-3. The number beside each reaction step corresponds to a numbered heading in the text.
Steps 1, 3, and 4 are essentially irreversible in the cell; all of the others are reversible.
Steps in the oxidative decarboxylation of pyruvate to acetyl-CoA by the pyruvate dehydrogenase complex.
In step 1 pyruvate reacts with the bound thiamine pyrophosphate (TPP) of pyruvate dehydrogenase (E1), undergoing decarboxylation to form the hydroxyethyl derivate.
Pyruvate dehydrogenase also carries out step
2 , the transfer of two electrons and the acetyl group from TPP to the oxidized form of the lipoyllysyl group of the core enzyme, dihydrolipoyl transacetylase (E
2
), to form the acetyl thioester of the reduced lipoyl group.
Step 3 is a transesterification in which the -SH group of CoA replaces the -SH group of E
2 to yield acetyl-CoA and the fully reduced
(dithiol) form of the lipoyl group. In step 4 dihydrolipoyl dehydrogenase (E3) promotes transfer of two hydrogen atoms from the reduced lipoyl groups of E
2 to the FAD prosthetic group of E
3
, restoring the oxidized form of the lipoyllysyl group of E
2
(in bold box). In step 5 the reduced FADH
2
E
3 group on transfers a hydride ion to NAD + , forming
NADH. The enzyme complex is now ready for another catalytic cycle.
E
1
E
2
E
3
- pyruvate dehydrogenase
- dihydrolipoyl transacetylase
- dihydrolipoyl dehydrogenase
The noncyclic reactions that provide biosynthetic precursors in anaerobically growing bacteria. These cells lack
-ketoglutarate dehydrogenase and therefore cannot carry out the complete cytric acid cycle.
-
Ketoglutarate and succinyl-CoA serve as precursors in a variety of biosynthetic reactions.
Intermediates of the citric acid cycle are drawn off as precursors in many biosynthetic pathways , yielding the products in the shaded areas. Four anaplerotic reactions that replenish depleted intermediates of the cytric acid cycle (broken arrows).
Regulation of metabolite flow from pyruvate through the citric acid cycle.
The pyruvate dehydrogenase complex is allosterically inhibited at high [ATP]/[ADP], [NADH]/[NAD + ], and [acetyl-CoA]/[CoA] rations, all of which indicate the energy-sufficent metabolic state. When these rations decrease, allosteric activation of pyruvate oxidation results. The rate of flow through the citric acid cycle can be limited by the availability of the substrates oxaloacetate and acetyl-CoA or by the depletion of NAD + by its conversion to NADH, which slows the three oxidation steps for which NAD + is the cofactor. Feedback inhibition by succinyl-CoA, citrate, and ATP also slows the cycle by inhibiting early steps. In muscle tissue, Ca 2+ signals contraction and stimulates energyyielding metabolism to replace the ATP consumed by contraction.
A summary of the points of entry of the standart amino acids into the citric acid cycle.
Some amino acids are listed more than once; these are broken down to yield different fragments, each of which enters the citric acid cycle at a different point. This scheme represents the major catabolic pathways in vertebrate animals, but there are minor variations from organism to organism.
Biochemical anatomy of a mitochondrion.
The convolutions (cristae) of the inner membrane give it a very large surface area. The inner membrane of a single liver mitochondrion may have over 10 000 sets of electron transfer system
(respiratory chains) and ATP synthase molecules, distributed over the whole surface of the inner membrane. Heart mitochondria, which have very profuse cristae and thus a much larger area of inner membrane, contain over three times as many sets of electron transfer systems as liver mitochondria. The mitochondrial pool of coenzymes and intermediates is functionally separated from the cytosolic pool.
The mitochondria of invertebrates, plants, and microbial eukaryotes are similar to those shown here, although there is much variation in size, shape, and degree of convolution of the inner membrane.
Path of electrons from NADH, succinate, fatty acyl-CoA, and glycerol-3-phosphate to ubiquinone
(UQ).
Electrons from NADH pass through a flavoprotein to a series of iron-sulfur proteins (in Complex I ) and then to UQ. Electrons from succinate pass through a flavoprotein and several Fe-S centers (in Complex
II ) on the way to UQ. Glycerol-3phosphate donates electrons to a flavoprotein (glycerol-3-phosphate dehydrogenase) on the outer face of the inner mitochondrial membrane, from which they pass through Fe-S centers to UQ. Acyl-CoA dehydrogenase (the first enzyme in b oxidation) transfers electrons to electron-transferring flavoprotein
(ETFP), from which they pass via
ETFP-ubiquinone oxidoreductase to
UQ.
The path of electrons through the
Complex III probably involves a "Q cycle" such as that shown here (blue arrows). The broken arrows represent diffusion of UQH
2
(ubiquinol) or its oxidized form UQ across the membrane. Notice that the electron transfers between cytochromes and ubiquinone are one-electron reactions, producing the semiquinone radical as an intermediate. The net effects of the reactions here are movement of electrons from UQH
2 to cytochrome c, and movement of protons from the inside (matrix) to the outer (cytosolic) side of the inner membrane (the intermembrane space).
Path of electrons through Complex IV.
Cu
A
(Cu 2+ ) and cyt a (Fe 2+ ) from one bimetallic redox center capable of accepting two electrons; Cu
B and cyt a 3 constitute a second two-electron redox center. The detailed path of electron flow between cyt c and O
2 is not known with certainty; apparently electrons first move from cyt c to Cu
A or cyt a, which are in rapid redox equilibrium with each other.
This bimetallic center then donates electrons to Cu
B and cyt a 3, also in redox equilibrium, which in turn donate the electrons that reduce O
2 to H
2
O. The four protons used in the reduction of O
2 to
H
2
O are taken up from the matrix side of the inner mitochondrial membrane.
Consequently, cytochrom oxidase pumps protons out of the matrix as electrons are transferred to O
2
.
The ATP synthase complex from mitochondria.
(a) Electon micrographs the knoblike protrusions from the mitochondrial inner membrane.
(b) Schematic diagram showing the likely organization of the subunits to form the protonconducting F
0 portion, and the ATP-synthesizing F three
, and one each of
,
, and
subunits.
1 unit. The F
1 complex consists of three
,
Summary of the flow of electrons and protons through the four complexes of the respiratory chain.
Electrons reach UQ via Complexes I and II. UQH
2
(ubiquinol) serves as a mobile carrier of electrons and protons. It passes electrons to Complex III, which passes them to another mobile connecting link, cytochrome c . Complex IV transfers electrons from reduced cytochrome c to O
2
. Electron flow through Complexes I, III, and IV is accompanied by proton flow from the matrix to the intermembrane space. Recall that electrons from fatty acid b oxidation can also enter the respiratory chain through UQ.
In this simple version of the chemiosmotic theory applied to mitochondria, electrons from NADH and other oxidizable substrates pass through a chain of carriers (cytochromes, etc.) arranged asymmetrically in the membrane. Electron flow is accompanied by proton transfer across the mitochondrial membrane, producing both a chemical (
pH) and an electrical
(
) gradient . The inner mitochondrial membrane is impermeable to protons; protons can reenter the matrix only through proton-specific channels (F
0
). The proton-motive force that drives protons back into the matrix provides the energy for ATP synthesis, catalyzed by the F
1 complex associated with F
0
.
The malate-asparate shuttle for transporting reducing equivalents from cytosolic NADH into the mitochondrial matrix.
1 NADH in the cytosol (intermembrane space) passes two reducing equivalents to oxaloacetate, producing malate.
2 Malate is transported across the inner membrane by the malate-
-ketoglutarate transporter.
3 In the matrix, malate passes two reducing equivalents to NAD + ; the resulting matrix NADH is oxidized by the mitochondrial respiratory chain. The oxaloacetate formed from malate cannot pass directly into the cytosol. It is first transaminated to form asparate 4 , which can leave via the glutamate-aspartate transporter 5 . Oxaloacetate is regenerated in the cytosol 6 , completing the cycle.
The glycerol-3-phosphate shuttle , an alternative means of moving reducing equivalents from the cytosol to the mitochondrial matrix.
Dihydroxyacetone phosphate in the cytosol accepts two reducing equivalents from cytosolic NADH in a reaction catalyzed by cytosolic glycerol-3-phosphate dehydrogenase. A membrane-bound isozyme of glycerol-3-phosphate dehydrogenase, located on the outer face of the inner membrane, transfers two reducing equivalents from glycerol-3-phosphate in the intermembrane space to ubiquinone.
Note that this shuttle does not involve membrane transport systems.
The uncoupling protein ( thermogenin ) of brown fat mitochondria, by providing an alternative route for protons to reenter the mitochondrial matrix, causes the energy conserved by proton pumping to be dissipated as heat.
Interlocking regulation of glycosis, pyruvate oxidation, the citric acid cycle, and oxidative phosphorylation by the relative concentrations of ATP, ADP, and AMP, and by NADH . High [ATP] (or low [ADP] and [AMP]) produces low rates of glycolysis, pyruvate oxidation, acetate oxidation via the citric acid cycle, and oxidative phosphorylation. All four pathways are accelerated when there is an increase in the rate of ATP utilization and increased formation of ADP, AMP, and P i
. Interlocking of glycolysis and the citric acid cycle by citrate, which inhibits glycolysis, supplements the action of the adenine nucleotide system. In addition, increased levels of NADH and acetyl-CoA also inhibit the oxidation of pyruvate to acetyl-CoA, and high [NADH]/[NAD + ] ratios inhibit the dehydrogenase reactions of the citric acid cycle.
Protein components of the mitochondrial electron transfer chain
Enzyme complex*
Mass
(kDa)
Number of subunits
Prosthetic group(s)
I NADH
dehydrogenase 850 >25 FMN, Fe-S
II Succinate
dehydrogenase
III Ubiquinone-
cytochrome
c
oxidoreductase
Cytochrome
c
140
250
4
10
FAD, Fe-S
Hemes, Fe-S
13 1 Heme
IV Cytochrome
oxidase
160 6-13 Hemes; Cu
A
, Cu
B
Sources : DePierre, J.W. & Ernster, L. (1977) Enzyme topology of intracellular membranes. Annu. Rev. Biochem . 46 , 201-262; Hafeti, Y. (1985) The mitochondrial electron transport and oxidative phosphorylation system. Annu. Rev.
Biochem . 54 , 1015-1069.
* Note that cytochrome c is not part of an enzyme complex, but moves between
Complexes III and IV as a freely soluble protein.
The stoichiometry of coenzyme reduction and ATP formation in the aerobic oxidation of a molecule of glucose via glycolysis, the pyruvate dehydrogenase reaction, and the citric acid cycle
Reaction
Number of ATP or reduced coenzymes directly formed
Number of ATP ultimately formed*
Glucose glucose-6-hosphate -1 ATP -1
Fructose-6-phosphate
fructose-1,6-bisphosphate
-1 ATP -1
2 Glyceraldehyde-3-phosphate
2 1,3-bisphosphoglycerate
2 1,3-Bisphosphoglycerate
2 3-phosphoglycerate
2 Phosphoenolpyruvate
2 pyruvate
2 Pyruvate 2 acetyl-CoA
2 NADH
2 ATP
2 ATP
2 NADH
6
2
2
6
2 Isocitrate
2
-ketoglutarate
2
-Ketoglutarate
2 succinyl-CoA
2 Succinyl-CoA 2 succinate
2 Succinate
2 fumarate
2 NADH
2 NADH
2 ATP (or 2
GTP)
2 FADH
2
6
6
2
4
2 Malate 2 oxaloacetate 2 NADH 6
Total 38
* This is calculated as 3 ATP per NADH and 2 per ATP per FADH
2
. A negative value indicates consumption.
http://www.brookscole.com/chemistry_d/templates/student_resources/shared_resources/animations/oxidativ e/oxidativephosphorylation.html