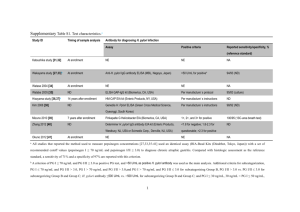
H. pylori
advertisement

Another YCP controversy: Membrane Potential influence with Clinical index and Bismuth Subcitrate Rol in some Chileans Study. Flow Cytometry viability analisys and Real Clinical Impact from this new way to evaluate Bismuth antibacterian actions. • Correlation between Flow Cytometry in the evaluation of Membrane Potential and Cellular Viability from HP • And Actual Rol from Bismuth Subcitrate in relation to the membrane equilibrium and Clinical Impact Departamento de Microbiología Facultad de Ciencias Biológicas Universidad de Concepción CITOMETRÍA DE FLUJO EN LA EVALUACIÓN DE POTENCIAL DE MEMBRANA Y VIABILIDAD CELULAR DE Helicobacter pylori. Magíster T.M. Juan Luis Castillo Navarrete Título Citometría de flujo en la evaluación de potencial de membrana y viabilidad celular de Helicobacter pylori. • Magister Juan Luis Castillo, Dra Apolinaria Garcia, Dr, Fernando Kawaguchi, Dr Carlos Gonzalez, Dr Jaime Madariaga, Sr, Ignacio Alfaro, Tamara Perez, Cinthya Perez. • Departamento de Microbiología, Facultad de Ciencias Biológicas, Universidad de Concepción. • Laboratorio de Citometría de Flujo, Hospital del Trabajador Concepción. Helicobacter pylori Patógeno gastroduodenal humano Hábitat pH óptimo Mucosa gástrica Agente etiológico: 6.0 - 7.0 (neutrófila) Ureasa • • • • Enfermedad péptica ulcerosa Dispepsia no ulcerosa Gastritis crónica superficial Linfoma MALT Crecimiento a pH < 4.5 Gastroenterology 1992: 102;720-727 Helicobacter pylori Estómago Capacidad de generar potencial bioenergético • Viabilidad • Crecimiento • Proliferación Potencial de membrana Gradiente electroquímico de protones (membrana interna) Bacterias aeróbicas • Egreso de protones • Ingreso de electrones Ingreso / Egreso de solutos Interior negativo -100 a -200 mV Fuerza protón motriz Gradiente eléctrico Potencial de membrana J Exp Biol 2000:203;51-59 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Potencial de membrana Disminución Involucrado Aumento Depolarización • Autolisis bacteriana • Transporte de glucosa • Quimiotaxis Hiperpolarización Se reduce a cero Ionóforos J Exp Biol 2000:203;51-59 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Impermeabilidad membrana celular Sustancias fluorescentes / vitales Indicador de viabilidad celular • Compuestos orgánicos con al menos 2 cargas (+) • Compuestos cargados negativamente Excluidos por membranas celulares intactas (eucariontes y procariontes) Letter Appl Microbiol 2002:34;1-7 FEMS Microbiol Rev 2000:24;429-448 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Membrana Celular Potencial de membrana Impermeabilidad de la membrana a ciertas sustancias Indicador de estado fisiológico de la membrana Viabilidad celular Letter Appl Microbiol 2002:34;1-7 FEMS Microbiol Rev 2000:24;429-448 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Permeabilidad membrana celular Indicador de muerte celular Compuestos fluorescentes que se unen a ácidos nucleicos • • • • • Propidium iodado (PI) TO-PRO-3 TO-PRO-1 Sytox Green Bromuro de etidio PI Viabilidad celular Letter Appl Microbiol 2002:34;1-7 FEMS Microbiol Rev 2000:24;429-448 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Potencial de membrana Eucariontes Procariontes Citometría de Flujo Bacterias en crecimiento activo Microelectrodos Cationes lipofílicos radiomarcados Valor promedio de suspensión celular completa J Exp Biol 2000:203;51-59 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Compuestos para ensayos Potenciométricos DiOC2(3) (488 nm) Carbocianinas (slow response probes) •Fluorescencia verde: • Se altera con el tamaño celular y presencia de agregados. • Es independiente del PM • Fluorescencia roja: • Dependiente del tamaño y del PM •Razón F. roja / F. verde: • Medida de PM Independiente del tamaño Potencial de Membrana: Indicador confiable de muerte celular DiOC2(3): (3,3´-dietiloxacarbocianina iodada Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 Handbook of fluorescent probes, 2002. (www.probes.com) Objetivos Generales Determinar por citometría de flujo, los parámetros de tamaño, forma celular, viabilidad celular y potencial de membrana en bacterias. Determinar el efecto que ejercen sobre el potencial de membrana y la viabilidad celular de H. pylori, diversas sustancias que tienen acción contra esta bacteria. Objetivos específicos Determinar, mediante citometría de flujo, los parámetros de tamaño, forma, viabilidad celular y potencial de membrana, en E. coli y S. aureus. Determinar, mediante citometría de flujo, los parámetros de tamaño, forma, viabilidad celular y potencial de membrana, en H. pylori. Evaluar el efecto de agentes antibacterianos usados comúnmente en el tratamiento de erradicación de H. pylori, sobre el potencial de membrana y viabilidad celular de H. pylori. Hipótesis Mediante citometría de flujo, es posible determinar los parámetros de tamaño, forma celular, potencial de membrana y viabilidad celular en Helicobacter pylori. Los diversos agentes antibacterianos utilizados en el tratamiento de la infección por Helicobacter pylori, producen alteraciones en la células bacteriana, tanto de tamaño, morfología, como también en el potencial de membrana y viabilidad celular; parámetros determinables mediante citometría de flujo. Estimación de bio volumen bacteriano DY en Gram positivos y Gram negativos Ácidos nucleicos y viabilidad celular Efecto de antimicrobianos sobre bacterias. Análisis monoparamétrico de poblaciones celulares Cytometry 2001:44;188-94 Appl Environ Microbiol 1998:64;3900-9 Cytometry 2000:41;41-5 Cytometry 1999:35;55-63 Antimicrob Agents Chemother 2000:44;827-834 Methods 2000:21;271-279 Cytometry 2001:43;223-226 Antimicrob Agents Chemother 2000:44;682-687 J Microbiol Methods 2000:42(1);1-2 Letters Appl Microbiol 2002:34;1-7 Cytometry 1997:29;298-305 Cytometry 1994:17;302-309 Cytometry 1994:17;302-9 Antimicrob Agents Chemother 2000:44;676-681 Antimicrob Agents Chemother 1998:42;1195-1199 FEMS Microbiol Rev 2000:24;429-448 Antimicrob Agents Chemother 2000:44;682-687 Cytometry Part A 2003:53A;97-102 Cytometry 1998:32;241-254 Cytometry 2001:43;55-68 Cepas bacterianas Escherichia coli ATCC 25922 Staphylococcus aureus ATCC 29213 Helicobacter pylori ATCC 43504 Escherichia coli K12 Pseudomona aeruginosa Acinetobacter spp. Bacillus subtilis Helicobacter pylori 636-C, 594-C, 612-C, 632-C633-A Viabilidad celular Potencial de membrana Eflujo a BrEt Estimación de patrones de tamaño bacteriano Cultivo líquido (18 hrs, 37°C) en caldo Müller Hinton filtrado (0.20 mm) Bacterias vivas Bacterias muertas Protocolo básico de trabajo Cultivo líquido (18 hrs, 37°C) Tubo Células vivas (ml) Células muertas (ml) 1 15 0 2 12 3 3 9 6 4 6 9 5 3 12 6 0 15 Protocolo de evaluación de viabilidad • 15 ml células vivas y/o muertas • 1.0 ml de PBS • 10 ml PI (1 mg/ml) • Incubar 5 minutos a T° ambiente y en oscuridad • Adquirir en Citómetro de flujo • Analizar fluorescencia roja Protocolo de evaluación de DY • 15 ml células vivas y/o muertas • 1.0 ml de PBS (o PBS-EDTA) • 30 ml DiOC2(3) 1 mM (30 mM Concentración final) • Incubar 4 minutos a T° ambiente y en oscuridad • Adquirir en Citómetro de flujo • Analizar fluorescencia roja y verde Metodología Citometría de Flujo Equipo • FACSCalibur, B.D. • Láser de Argón, 488 nm. Adquisición • Software CellQuest v3.3 • Máximo de 1000 eventos/seg. • 20.000 eventos celulares Análisis • Software CellQuest v.3.3 • Media de intensidad de fluorescencia verde y roja. • Cálculo razón F. roja / F. verde • Software Flow Explorer Rev Méd Chile 1999:127;1385-1397 Antimicrob Agents Chemother 2000:44;827-834 Cytometry 1999:35;55-63 / 2001:43;223-226 http://www.cyto.purdue.edu/flowcyt/software/flowex4_files/Install4.exe Tubo 1PM.001 PM.001 (CELLQuest) Email Flow explorer CELLQuest Cáculo de razón FL2/FL1 FACS Convert Nueva fila (1pm.001) Gráficos Mac Email Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori Definición de gate de análisis Estimación de patrones de tamaño Markers (+/- channels) M1 M2 M3 P. aeuriginosa Acinetobacter sp. M4 M5 M6 M7 M8 E. coli B. subtilis Estimación de patrones de tamaño Estimación de patrones de tamaño en cepas de H. pylori Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori Incorporación de PI Células vivas Células vivas y muertas (1:1) 51.37% 0.98% Células muertas 99.03% E. coli:E.coli Incorporación dede PI porcélulas células muertas por calor : Incorporación PI por muertas 120,00 100,00 Porcentaje R = 0.997 80,00 60,00 40,00 20,00 0,00 0,000 3,000 6,000 9,000 12,000 15,000 Células muertas x 10 6 S. aureus: Incorporación PIpor por células muertas por calor S. aureus: Incorporaciónde de PI células muertas 90,00 80,00 R = 0.692 Porcentaje 70,00 60,00 50,00 40,00 30,00 20,00 10,00 0,00 0,000 3,000 6,000 9,000 Células muertas x 10 12,000 4 15,000 Agregados y destrucción celular E.coli : Incorporación de por PI por células muertas muertas (shock frío). E. coli: Incorporación de PI células (Shock frío). Ensayo por triplicado. Ensayo en triplicado. 100,00 90,00 R = 0.978 80,00 Porcentaje 70,00 60,00 50,00 40,00 30,00 20,00 10,00 0,00 0,000 3,000 6,000 9,000 Células muertas x 10 12,000 15,000 6 S. aureus : Incorporación depor PI por célulasmuertas muertas (shock frío). S. aureus: Incorporación de PI células (Shock frío). Ensayo por triplicado. Ensayo en triplicado. 100,00 90,00 R = 0.981 80,00 Porcentaje 70,00 60,00 50,00 40,00 30,00 20,00 10,00 0,00 0,000 3,000 6,000 9,000 Células muertas x 104 12,000 15,000 Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori Estimación de DY dep. pol. Células vivas dep. dep. pol. Células vivas y muertas (1:1) pol. Células muertas Estimación de DY dep. dep. Flow pol. Explorer pol. Razón FL2/FL1 Células vivas y muertas (1:1) S. aureus: Células polarizadas y depolarizadas. Razón flourescencia roja / verde (DioC2(3)). Ensayo por triplicado. 100,00 R = - 0.982 (pol) 90,00 80,00 Porcentaje 70,00 60,00 50,00 40,00 30,00 20,00 R = 0.982 (dep) 10,00 0,00 0,000 3,000 6,000 9,000 12,000 15,000 Células muertas x 10 4 E. coli: Células polarizadas y depolarizadas. Razón flourescencia roja / verde (DioC2(3)). Ensayo por triplicado. 100,00 90,00 R = - 0.993 (pol) 80,00 Porcentaje 70,00 60,00 50,00 40,00 30,00 20,00 R = 0.993 (dep) 10,00 0,00 0,000 3,000 6,000 9,000 Células muertas x 10 12,000 6 15,000 Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori H. pylori: Incorporación de PI H. pylori: Incorporación de DioC2(3) Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori Eflujo a Bromuro de Etidio Ingreso por difusión pasiva Intercalante Monocatiónico Sistema de eflujo del tipo MFS (Major facilitator superfamiliy): • Dependiente de la fuerza protón motriz • Sustratos: • Fluoroquinolonas hidrofílicas • Compuestos orgánicos monocatiónicos (Acriflavina, BrEt, Bromuro de tetrafenilfosfonio) Fluorescencia Balance entre influjo y eflujo Exclusión Bajo influjo y/o presencia de eflujo Protocolo de evaluación de eflujo a Bromuro de Etidio • 15 ml células vivas y/o muertas • 1.0 ml de PBS • 20 ml BrEt (20 mg/ml concentración final) • Incubar 5 minutos a T° ambiente y en oscuridad • Adquirir en Citómetro de flujo • Analizar fluorescencia roja. Cytometry 1994:17;302-309 Eflujo a Bromuro de Etidio 1.29 % Células vivas 97.95 % Células muertas Incorporación de PI y eflujo a bromuro de etidio E. coli S. auerus R=0.999 R=0.999 R=0.999 R=0.998 Resultados • Estimación patrones de tamaño • Incorporación de PI en S. aureus y E. coli • DY en S. aureus y E. coli • Incorporación de PI y DY en H. pylori • Eflujo a bromuro de etidio en S. aureus y E. coli • Eflujo a bromuro de etidio en H. pylori H. pylori: Eflujo a bromuro de etidio H. pylori: Incorporación de PI H. pylori: Eflujo a BrEt H. pylori: Incorporación de DioC2(3) Discusión • Desarrollo de método de estimación de patrones de tamaño • Incorporación de PI en E.coli y S.aureus • Ensayo de DY en E.coli y S.aureus • Uso de Razón de Fluorescencia Roja/verde • H. pylori: PI, BrEt y DY Discusión Desarrollo de método de estimación de patrones de tamaño 1. 2. 3. 4. Medición de FSC usada ampliamente Indice de refracción Aplicaciones Limitaciones: • Células en suspensión • Tamaño no cuantificable • Mezcla de poblaciones celulares 5. Ventajas: • Método sencillo y económico • Uso de parámetros intrínsecos a la células Discusión Incorporación de PI en E.coli y S.aureus 1. 2. 3. 4. 5. Concepto Controles Linealidad Definición de viabilidad celular Ventajas: • Método sencillo y económico • Combinación con parámetros intrínsecos a la células Discusión Ensayo de DY en E.coli y S.aureus 1. Metodología ampliamente establecida 2. Disponibilidad de inmunerables flurocromos (http://probes.invitrogen.com/handbook/sections/2200.html) 3. Aplicaciones 4. Limitaciones: • Células en suspensión y no aisladas • Costo, infraestructura y softwares de apoyo 5. Ventajas: • Método rápido • Información sobre poblaciones celulares • Posibilidad de cuantificar Discusión * *** DIOC2(3) Agregados de DIOC2(3) Fluorescencia verde Fluorescencia roja Razón Fluorescencia Roja / verde DY - -* - Uso de Razón de Fluorescencia Roja/verde = - ** *** ** *- -* *- DY - - - - * * * * * *- *** ***** **** *** *-* * -* ** ** ** **- *- -* < <<< = Proporcional al tamaño Sensible a presencia de agregados Sensible a DY e independiente del tamaño Discusión H. pylori: PI, BrEt y DY 1. Barrera de permeabilidad 2. Sistema de eflujo (operón hefABC: Nº acceso AF059041) 3. pH, medio ambiente y DY: • Adecuación del DY • DY positivo: • Uso de oxonoles (DiBAC4(3)) (evidencias células depolarizadas) 4. Características fisiológicas aún desconocidas Conclusiones 1 Se desarrolla un método de análisis que permite la estimación de patrones de tamaño celular de células bacterianas por citometría de flujo. 2 En E. coli y S. aureus, la citometría de flujo evalúa exitosamente viabilidad celular mediante la incorporación de yoduro de propidio, DY, usando DIOC2(3) y la presencia de eflujo a bromuro de etidio. Conclusiones 3 En E. coli y S. aureus, la evaluación por citometría de flujo de la incorporación de yoduro de propidio, la estimación de DY y la presencia de eflujo a bromuro de etidio, en conjunto se pueden considerar criterios para definir viabilidad celular 4 En H. pylori, bajo las condiciones experimentales ensayadas no fue factible evaluar viabilidad celular, definida por la incorporación de yoduro de propidio, DY, usando DIOC2(3) ni evaluar la presencia de eflujo a bromuro de etidio. Proyecciones Patrones de tamaño celular Estudio de acción de diversas sustancias sobre bacterias Incorporación de PI Viabilidad v/s cultivabilidad Estimación de DY Eflujo a BrEt Búsqueda de otras condiciones metodológicas en H. pylori Proyecciones Este trabajo sienta las bases para el desarrollo de una línea de trabajo relacionada con la presencia y actividad de sistemas de eflujo en bacterias, desde un punto de vista fisiológico y funcional. Estudio de sistemas de eflujo y resistencia a drogas en células eucariontes, en especial, células neoplásicas y el fenotipo MDR (Multi Drug Resistant), particularmente la presencia y actividad de eflujo de la glicoproteína p (DIOC2(3) es sustrato de la glicoproteína p). El estudio integral de viabilidad celular en E. coli y S. aureus, es hasta donde se tiene conocimiento, pionero en nuestro país, por lo que debiera fomentar el uso de la citometría de flujo en estudios de microbiológicos y en especial bacteriológicos. • Similar morphologic and outer membrane changes were observed following growth in ironlimiting medium and at the MICCBS that inhibited the growth of all three strains. • These changes, which were also observed for iron-limited bacteria, were alleviated by the addition of iron to the cultures. • H. pylori ATP levels, reduced in iron-limiting medium, were below the limits of detection in two of the three strains following exposure to bismuth. • The addition of iron partially restored bacterial ATP levels in these two strains, although not to normal concentrations. • In contrast, exposure of the same strains to the MICCBS failed to deplete intracellular levels of iron, which were significantly reduced by culturing in iron-limiting medium. • Thus, the antimicrobial effect of bismuth and of iron limitation on H. pylori may be similar. • However, the respective mechanisms of intracellular action would appear to be mediated by different pathways within the cell. • Prior to the discovery of the link between Helicobacter pylori and peptic ulcer disease (10), ulcers were thought to result from abnormal levels of gastric acid secretion. • Bismuth compounds, commonly prescribed for ulcers, were initially thought to act as a barrier to the digestive effects of stomach acid by coating the ulcer site (31). • Instead, it appears that H. pylori is highly susceptible to bismuth compounds (39), and treatment with colloidal bismuth subcitrate (CBS) is associated with a reduction in bacterial numbers and a concurrent reduction in gastritis in vivo (37). • Despite these findings, bismuth monotherapy often fails to completely eradicate these bacteria (13, 43), an observation that correlates with increasing ulcer relapse rates over time (34). • The oldest regimen, an extremely effective one for clearing H. pylori infection, uses bismuth in combination with certain antibiotics (2, 51). • Bismuth “triple therapy” consists of a bismuth compound (usually CBS or bismuth subsalicylate) in combination with metronidazole and tetracycline or amoxicillin (16) and reportedly cures 87.9% of patients within 1 week of treatment and 89.2% of patients within 2 weeks of treatment (49) • H. pylori strains resistant to bismuth have not been reported and presumably arise at a lower frequency than strains resistant to antimicrobial agents such as nitroimidazoles, macrolides, and tetracycline (40). • Bismuth compounds may reduce the development of resistance to coadministered antibiotics (27) and are also effective at treating H. pylori strains with established resistance to other antibiotics (3, 40). • How bismuth is toxic to H. pylori is not known. • A number of studies have linked the antimicrobial activities of many heavy metals, including bismuth, to their effects on iron uptake by bacteria (1, 21, 23, 24). • Iron is required for growth because it is a cofactor for many essential enzymatic processes (42), and many of the observed effects of bismuth on bacteria could be the result of iron limitation. • These effects include a reduction in intracellular ATP levels (44), inhibition of protein and cell wall synthesis because loss Sodio/Potasio Bumb equilibrium of membrane potential function (32) with reduction in capsular polysaccharide. • Prior to the discovery of the link between Helicobacter pylori and peptic ulcer disease (10), ulcers were thought to result from abnormal levels of gastric acid secretion. • Bismuth compounds, commonly prescribed for ulcers, were initially thought to act as a barrier to the digestive effects of stomach acid by coating the ulcer site (31). • Instead, it appears that H. pylori is highly susceptible to bismuth compounds (39), and treatment with colloidal bismuth subcitrate (CBS) is associated with a reduction in bacterial numbers and a concurrent reduction in gastritis in vivo (37). • Despite these findings, bismuth monotherapy often fails to completely eradicate these bacteria (13, 43), an observation that correlates with increasing ulcer relapse rates over time (34). • The oldest regimen, an extremely effective one for clearing H. pylori infection, uses bismuth in combination with certain antibiotics (2, 51). • Bismuth “triple therapy” consists of a bismuth compound (usually CBS or bismuth subsalicylate) in combination with metronidazole and tetracycline or amoxicillin (16) and reportedly cures 87.9% of patients within 1 week of treatment and 89.2% of patients within 2 weeks of treatment (49). • H. pylori strains resistant to bismuth have not been reported and presumably arise a lower frequency than strains resistant to antimicrobial agents such as nitroimidazol macrolides, and tetracycline (40). • Bismuth compounds may reduce the development of resistance to coadministered antibiotics (27) and are also effective at treating H. pylori strains with established resistance to other antibiotics (3, 40). • Even this mechanism is clear, habitually gastroenterologists don’t know how bismut toxic to H. pylori is not known. • A number of studies have linked the antimicrobial activities of many heavy metals, including bismuth, to their effects on iron uptake by bacteria (1, 21, 23, 24). • Iron is required for growth because it is a cofactor for many essential enzymatic processes (42), and many of the observed effects of bismuth on bacteria could be the result of iron limitation. • These effects include a reduction in intracellular ATP levels (44), inhibition of prot and cell wall synthesis and of membrane function (32), and a reduction in capsular polysaccharide production (19, 20, 22, 28). • A reduction in outer membrane lipopolysaccharide (LPS) expression occurs when H. pylori is deprived of iron (J. Keenan, unpublished data). Castillo’s group already demonstrated through flow cytometry, celular form, size, membrane potential and celular viability. Again, Several iron-repressible outer membrane proteins are concomitantly up-regulated (18, 29, 52). • These observations led us to hypothesize that the antimicrobial action of bismuth could be due to it competitively inhibiting iron uptake. • To test this hypothesis, we searched for indicators of iron limitation in bismuth-exposed H. pylori and investigated whether iron could protect these bacteria from the antimicrobial effect of bismuth. • MATERIALS AND METHODS • Bacterial strains and cultures. Three well-characterized H. pylori strains were used in this study: H. pylori 60190 (ATCC 49503), a cag pathogenicity island (PAI)-positive toxigenic strain (5, 14); cag PAI-negative H. pylori strain Tx-30a (ATCC 51932) (14), which fails to produce detectable cytotoxin activity in vitro (35); and mouse-adapted H. pylori Sydney strain 1 (SS1) (33). Individual strains were grown in the base medium, brucella broth (BB; Difco, Detroit, Mich.) supplemented with 5% fetal bovine serum (FBS; Gibco BRL, Auckland, New Zealand), for 72 h at 37°C under microaerophilic conditions and with constant rotation (120 rpm). • • Iron limitation and bismuth inhibition were achieved by growing the strains in BB-5% FBS with the addition of an iron chelator (deferoxamine mesylate; Sigma, St. Louis, Mo.) at a final concentration of 50 μM or CBS at the MIC (MICCBS) determined for each strain (see below). • Iron salts (ferrous ammonium sulfate; Sigma) at final concentrations of 50 to 1,000 μM were added to the base medium containing MICCBS to determine the amount of iron required to alleviate the effects of bismuth. • The deferoxamine, CBS, and iron salts solutions were prepared immediately before use. Deferoxamine and iron salts solutions were filter sterilized prior to addition to the medium.Morphologic changes. • The morphologic changes in the bacteria were assessed by transmission electron microscopy (TEM) over the 72-h r period. Washed bacteria were placed on carbon-colloidin-coated mesh grids and negatively stained with 1% aqueous phosphotungstic acid (pH 7.0). • Photographs were taken by using a CM12 transmission electron microscope (Philips).Determining MICCBS values for individual H. pylori strains. • MICCBS values were determined by using 12-well tissue culture plates inoculated with CBS stock solution to a final volume of 2 ml/well. • CBS, prepared from bismuth citrate powder and ammonia (26), was combined with 2 × 107 H. pylori organisms per ml at final concentrations of 0, 1, 2, 4, 8, 16, 32, and 64 mg/liter in the base medium, and the mixture was incubated for 72 h in a microaerophilic environment with constant rotation. • H. pylori growth assessed by measuring the optical density at 650 nm in each well (corrected by using uninoculated controls) was confirmed as the growth of gram-negative, urease-positive colonies on blood agar plates. • Each strain was tested six times per experiment, performed on three separate occasions, and controls containing ammonia without bismuth were included to ensure that any inhibitory effects were due to bismuth alone.OMV. • Bacteria were removed from 72-h broth cultures by centrifugation (10,000 × g, 15 min, 4°C), and the supernatants were ultracentrifuged (100,000 × g, 2 h, 4°C) to recover outer membrane vesicles (OMV) as described previously (30). • The OMV pellet was washed twice with phosphate-buffered saline (PBS) and assayed for protein content (36). OMV components separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis under reducing conditions were silver stained to visualize protein and LPS profiles (30). • OMV-associated protease activity was detected by zymography (29). • Briefly, OMV were electrophoresed under nonreducing conditions through an acrylamide gel containing a copolymerized substrate (gelatin). • After extensive washing to remove sodium dodecyl sulfate, the gel was incubated in 50 mM Tris buffer (pH 7.4) with 10 mM calcium chloride for 4 h at 37°C. • OMV-associated proteolytic activity was visualized as clear bands (indicative of substrate lysis) against the blue background of the gels following Coomassie blue staining.Collection of samples for analysis of ATP and intracellular iron levels. • H. pylori organisms were grown in BB-5% FBS for 24 h before being adjusted to the conditions described above for the remaining 48 h of culturing. • At 72 h, a 1-ml aliquot was removed to determine the effects of the various culture conditions on bacterial numbers by viable colony counting. A second aliquot was snap-frozen (in liquid nitrogen) and stored at −80°C prior to analysis of ATP levels. • The remaining bacteria were harvested (10,000 × g, 20 min) to determine intracellular iron levels. ATP and iron levels were determined in relation to bacterial dry weight.Viable colony counting. • After serial dilution in PBS, bacterial titers were inferred from CFU per milliliter, determined by spreading bacterial suspensions over the surface of blood agar plates containing 5% defibrinated sheep blood. • The agar plates were incubated for 5 days at 37°C under microaerophilic conditions (as described above).Analysis of ATP levels. • ATP levels were measured by using a luminescent ATP detection assay kit (ATPLite-Packard Bioscience) according to manufacturer directions. • Briefly, frozen samples were quickly defrosted, and 100-μl portions of the samples were added to the wells of 96-well tissue culture plates in duplicate. • Following the addition of 50 μl of lysis solution, the plates were shaken for 5 min (700 rpm), and then 50 μl of substrate buffer solution was added. • The plates were shaken for a further 5 min and dark adapted for 10 min before luminescence was measured with a luminometer (BMG Labtechnologies). • ATP standards, prepared for each of the four conditions used to culture H. pylori and ranging from 10−5 M to the blank, were used to generate a standard curve (Graphpad-Instat) from which unknown sample concentrations were calculated.Analysis of iron levels. • PBS-washed bacterial pellets were oven dried at 80°C for approximately 18 h in 1.5ml Eppendorf tubes and then transferred to 10-ml acid-washed flasks for the determination of dry weights. • Each sample was dissolved in 1 ml of concentrated nitric acid at 55 to 65°C overnight. • Dissolved samples were cooled to an ambient temperature and then brought to 10 ml with Milli-Q distilled water. • Samples were examined on a Spectra-10 (Varian) atomic absorption spectrometer, and iron concentrations were determined from a standard curve that ranged from 0 to 5,000 μg/liter (25). • Statistical analyses. The effects of each of the growth conditions on bacterial iron and ATP levels were evaluated by using a factorial analysis of variance. • Significant effects indicated by this analysis were further explored by using Fisher's least-significant-difference test. • The ATP measurements were natural logarithm transformed prior to analysis, so that very small values could be easily analyzed. • RESULTS • • • • • • • Bismuth or iron limitation induces a change in H. pylori morphology and outer membrane composition. The MICCBS values were found to be 4 mg/liter (10 μM) for strains 60190 and SS1 and 8 mg/liter (20 μM) for Tx-30a. The addition of ammonia alone (see Materials and Methods) did not inhibit the growth of any of the strains. Thus, the inhibitory effects of CBS were due to bismuth alone (results not shown). Each of the three H. pylori strains was examined by TEM following 72 h of growth in base medium (control), iron-limiting medium (containing 50 μM deferoxamine), or medium containing MICCBS (Fig. 1). Both iron limitation and exposure to MICCBS resulted in the transition of H. pylori from helical to coccoid morphology (Fig. 1A); a larger proportion of MICCBS-exposed bacteria had completed this transition by 72 h. Exposure to MICCBS resulted in some more distorted forms of H. pylori (Fig. 1B). In contrast, bacteria grown under normal culture conditions exhibited normal curved-rod morphology. • • RESULTS Bismuth or iron limitation induces a change in H. pylori morphology and outer membrane composition. • The MICCBS values were found to be 4 mg/liter (10 μM) for strains 60190 and SS1 and 8 mg/liter (20 μM) for Tx30a. • The addition of ammonia alone (see Materials and Methods) did not inhibit the growth of any of the strains. • Thus, the inhibitory effects of CBS were due to bismuth alone (results not shown). • Each of the three H. pylori strains was examined by TEM following 72 h of growth in base medium (control), ironlimiting medium (containing 50 μM deferoxamine), or medium containing MICCBS (Fig. 1). • • Both iron limitation and exposure to MICCBS resulted in the transition of H. pylori from helical to coccoid morphology (Fig. 1A) A larger proportion of MICCBS-exposed bacteria had completed this transition by 72 h. Exposure to MICCBS resulted in some more distorted forms of H. pylori (Fig. 1B). • In contrast, bacteria grown under normal culture conditions exhibited normal curved-rod morphology • There was little observable difference in the outer membrane proteins of individual • H. pylori strains following iron limitation or exposure to bismuth (Fig. 2). • Specific markers for iron stress were difficult to identify, but a 12-kDa protein band was clearly enhanced in the presence of MICCBS or 50 μM deferoxamine (Fig. 2) • . Both iron-limited and MICCBS-exposed bacteria also had reduced levels of OMV-associated LPS (Fig. 3). • In addition, zymography revealed OMV-associated protease activity in each strain following growth in the presence of MICCBS or under iron-limiting conditions (Fig. 4). • Iron protects H. pylori from the inhibitory effects of bismuth. • The addition of 250 μM iron protected strains 60190 and SS1 from the inhibitory effects of 10 μM CBS. H. pylori Tx-30a, which required twofold more bismuth for growth inhibition (20 μM), also required more iron (500 μM) to achieve any protective effect. • However, this amount of iron only reduced the bactericidal effect of bismuth on Tx-30a, as indicated by growth after culturing on blood agar. • It did not completely reverse growth inhibition, as demonstrated by a slight increase in absorbance readings after 72 h of culturing. • H. pylori grown in medium containing MICCBS and protective iron maintained the normal curved-rod morphology typical of these bacteria (Fig. 1D). • Furthermore, this treatment of H. pylori 60190 (results not shown) and SS1 (Fig. 2) resulted in outer membrane protein profiles similar to those of bacteria grown in base medium. • The provision of protective iron to H. pylori Tx-30a apparently failed to decrease the expression of the 12-kDa protein band (Fig. 2B). • We speculate that this result may have been due to the visible overloading of the corresponding lane. • However, it is possible that this protein is still expressed at a higher level in this strain under these conditions. • Protease activity was no longer detectable in bacteria of all three strains grown in medium containing MICCBS and protective iron (Fig. 4) • ATP but not iron levels are reduced following exposure of H. pylori to bismuth. • Intracellular iron levels of H. pylori exposed to MICCBS were compared with those of bacteria cultured under normal and iron-limiting conditions to assess whether bismuth inhibited iron uptake. To achieve the biomass required for this analysis, cultures were grown overnight before being exposed to bismuth or limiting iron for 48 h. • • Colony counts confirmed that the established MICCBS values were sufficient to prevent growth even at these much higher concentrations of bacteria, and inhibition was still overcome by the addition of iron at protective concentrations (results not shown). • PBS used to wash the bacterial pellet was assayed for iron by atomic absorption spectrometry. • After three washes, the amount of iron present in the PBS supernatant was below the detectable limits of the assay, indicating this number of washes to be sufficient for removing excess unbound iron (results not shown). • H. pylori strains 60190 and SS1 grown in the presence of MICCBS had intracellular levels of iron similar to those of bacteria cultured under normal conditions, as measured by atomic absorption spectrometry. • In contrast, iron levels were significantly reduced following culturing in 50 μM deferoxamine (Fig. 5). • However, MICCBS had an effect similar to that of iron limitation on the intracellular iron levels of H. pylori Tx-30a. • All three strains cultured in the presence of MICCBS and protective iron had levels of intracellular iron that were higher than those of controls but not significantly different from those of bacteria cultured with protective iron alone.Bacterial • ATP levels were only significantly reduced in strains SS1 and 60190 following growth in ironlimiting medium (Fig. 6). Exposure to MICCBS resulted in a reduction in ATP production to below the detectable limits of the assay for these two strains, whereas ATP levels in H. pylori Tx-30a were not significantly different from those in controls. • • • The addition of iron to bacterial cultures containing MICCBS increased bacterial ATP concentrations in all three strains. However, protective iron failed to restore H. pylori 60190 ATP levels to those observed following culturing under normal or even iron-limiting conditions • RESULTS • Bismuth or iron limitation induces a change in H. pylori morphology and outer membrane composition. • The MICCBS values were found to be 4 mg/liter (10 μM) for strains 60190 and SS1 and 8 mg/liter (20 μM) for Tx30a. • The addition of ammonia alone (see Materials and Methods) did not inhibit the growth of any of the strains. • Thus, the inhibitory effects of CBS were due to bismuth alone (results not shown). • Each of the three H. pylori strains was examined by TEM following 72 h of growth in base medium (control), ironlimiting medium (containing 50 μM deferoxamine), or medium containing MICCBS (Fig. 1). • Both iron limitation and exposure to MICCBS resulted in the transition of H. pylori from helical to coccoid morphology (Fig. 1A); a larger proportion of MICCBS-exposed bacteria had completed this transition by 72 h. Exposure to MICCBS resulted in some more distorted forms of H. pylori (Fig. 1B). • In contrast, bacteria grown under normal culture conditions exhibited normal curved-rod morphology.There was little observable difference in the outer membrane proteins of individual H. pylori strains following iron limitation or exposure to bismuth (Fig. 2). • Specific markers for iron stress were difficult to identify, but a 12-kDa protein band was clearly enhanced in the presence of MICCBS or 50 μM deferoxamine (Fig. 2). • Both iron-limited and MICCBS-exposed bacteria also had reduced levels of OMV-associated LPS (Fig. 3). In addition, zymography revealed OMV-associated protease activity in each strain following growth in the presence of MICCBS or under iron-limiting conditions (Fig. 4) • .Iron protects H. pylori from the inhibitory effects of bismuth. • The addition of 250 μM iron protected strains 60190 and SS1 from the inhibitory effects of 10 μM CBS. H. pylori Tx-30a, which required twofold more bismuth for growth inhibition (20 μM), also required more iron (500 μM) to achieve any protective effect. • However, this amount of iron only reduced the bactericidal effect of bismuth on Tx-30a, as indicated by growth after culturing on blood agar. • It did not completely reverse growth inhibition, as demonstrated by a slight increase in absorbance readings after 72 h of culturing. • H. pylori grown in medium containing MICCBS and protective iron maintained the normal curved-rod morphology typical of these bacteria (Fig. 1D). • Furthermore, this treatment of H. pylori 60190 (results not shown) and SS1 (Fig. 2) resulted in outer membrane protein profiles similar to those of bacteria grown in base medium. • The provision of protective iron to H. pylori Tx-30a apparently failed to decrease the expression of the 12-kDa protein band (Fig. 2B). • We speculate that this result may have been due to the visible overloading of the corresponding lane. • However, it is possible that this protein is still expressed at a higher level in this strain under these conditions • . Protease activity was no longer detectable in bacteria of all three strains grown in medium containing MICCBS and protective iron (Fig. 4) • ATP but not iron levels are reduced following exposure of H. pylori to bismuth. • Intracellular iron levels of H. pylori exposed to MICCBS were compared with those of bacteria cultured under normal and iron-limiting conditions to assess whether bismuth inhibited iron uptake. • To achieve the biomass required for this analysis, cultures were grown overnight before being exposed to bismuth or limiting iron for 48 h. • Colony counts confirmed that the established MICCBS values were sufficient to prevent growth even at these much higher concentrations of bacteria, and inhibition was still overcome by the addition of iron at protective concentrations (results not shown) • . PBS used to wash the bacterial pellet was assayed for iron by atomic absorption spectrometry. • After three washes, the amount of iron present in the PBS supernatant was below the detectable limits of the assay, indicating this number of washes to be sufficient for removing excess unbound iron (results not shown). • H. pylori strains 60190 and SS1 grown in the presence of MICCBS had intracellular levels of iron similar to those of bacteria cultured under normal conditions, as measured by atomic absorption spectrometry. In contrast, iron levels were significantly reduced following culturing in 50 μM deferoxamine (Fig. 5). • However, MICCBS had an effect similar to that of iron limitation on the intracellular iron levels of H. pylori Tx-30a. • All three strains cultured in the presence of MICCBS and protective iron had levels of intracellular iron that were higher than those of controls but not significantly different from those of bacteria cultured with protective iron alone.Bacterial ATP levels were only significantly reduced in strains SS1 and 60190 following growth in iron-limiting medium (Fig. 6). • Exposure to MICCBS resulted in a reduction in ATP production to below the detectable limits of the assay for these two strains, whereas ATP levels in H. pylori Tx-30a were not significantly different from those in controls. • • The addition of iron to bacterial cultures containing MICCBS increased bacterial ATP concentrations in all three strains. However, protective iron failed to restore H. pylori 60190 ATP levels to those observed following culturing under normal or even iron-limiting conditions • DISCUSSION • In this study, we have shown that exposure to MICCBS has the same effect on H. pylori as iron limitation during growth. H. pylori cultured in the presence of MICCBS underwent a morphologic conversion from the bacillary to the coccoid form that was associated with changes in outer membrane composition. • These changes, which were also observed in iron-limited bacteria, were prevented by supplementation of the medium with iron. • However, our hypothesis that the antimicrobial action of bismuth was due to the inhibition of iron uptake was challenged by unchanged intracellular iron levels in two of the three H. pylori strains following culturing in the presence of this bismuth compound.Bismuthinduced changes in H. pylori morphology have been seen before (11, 45, 47), but our observations of accompanying changes in outer membrane protein and LPS profiles are new. • The bismuth-induced changes included the up-regulation of a 12-kDa protein and two proteases and diminished LPS expression and are identical to those observed in other studies in which H. pylori was grown under iron-limiting conditions (9, 29). • Furthermore, the addition of iron during growth mostly prevented these bismuth-induced changes. One possible exception was the continued expression of a 12-kDa protein by H. pylori Tx-30a grown in the presence of MICCBS and protective iron, even though TEM revealed helical forms of bacteria under these conditions. • This observation may be linked to the incomplete reversal of bismuth-induced growth inhibition in the presence of protective iron. However, the apparent overloading of the corresponding lane may also be a contributing factor. The change from helical to coccoid morphology was more rapid in the bismuth-exposed bacteria and was associated with a significant decrease in viability. ATP levels were below the limits of detection in H. pylori strains 60190 and SS1 following exposure to bismuth, whereas strain Tx-30a had ATP levels not significantly different from those seen under normal culture conditions, despite a rapid conversion to the coccoid form that was likewise associated with decreased viability. • • Strain Tx-30a was also associated, again in contrast to the other two strains, with a significant reduction in intracellular iron levels following exposure to MICCBS. Both of these results would be consistent with a loss of membrane integrity (44), leading to an accumulation of nonutilizable ATP in the extracellular medium and a concurrent loss of iron from the cell. • We speculate that this effect may reflect the higher bismuth concentration needed to inhibit this strain and may also be linked to the incomplete reversal of bismuth-induced growth inhibition in the presence of protective iron.H. pylori 60190 and SS1 cultured with MICCBS and protective iron had ATP levels higher than those found in the presence of bismuth alone but significantly lower than those found under normal culture conditions. • • The use of protective iron and MICCBS was also associated with an increase in intracellular iron levels and partial protection from bismuthinduced growth inhibition. • These observations, which would suggest that bismuth and iron were competing for uptake via the same pathway in these bacteria, were further supported by our observation that iron limitation had a similar (but less severe) effect on H. pylori ATP levels and viability. • Unexpectedly, however, intracellular iron levels remained constant in these two strains following exposure to bismuth, whereas iron limitation over the same growth period resulted in the depletion of iron. • These results would suggest that the mechanism for the antimicrobial effect of bismuth on these and other bacteria is not simply iron starvation, as suggested previously (21). • Instead, the effect of bismuth only mimics the effect of iron starvation in H. pylori.ATP synthesis in bacteria occurs at the cytoplasmic membrane through the action of multisubunit enzymes (F1F0-ATPases) that utilize the membrane potential and electrochemical gradient generated by respiration (48). • In H. pylori, inhibition of these enzymes is thought to lead to ATP depletion and the associated inhibition of other pathways that are important for H. pylori survival (38). • Other potential targets for the inhibition of ATP synthesis include the iron-sulfur clusters and cysteine-containing heme groups that act as electron carriers in the respiratory chain. The observation that bismuth complexes localize in the periplasmic space (between the cytoplasmic and outer membranes) in H. pylori (4, 32) would be consistent with a mechanism of action of bismuth that may involve the inhibition of ATP synthesis via one or more of these pathways • The accumulation of bismuth at the cytoplasmic membrane might have prevented iron incorporation into iron-utilizing proteins, thereby creating a functional iron limitation within the respiratory chain. Thus, the bismuth-induced production of iron-repressible outer membrane proteins could be a response to perceived iron stress, despite no change in intracellular iron levels, an effect similar to that caused by other heavy metals through competition for incorporation into iron binding proteins (8). • However, bismuth is also able to inhibit the electron transport chain through binding to the sulfhydryl groups within the enzyme complex, and this mechanism might instead account for the rapid reduction in intracellular ATP levels (6, 7). • Interestingly, McGowan et al. showed that F1F0-ATPase activity is required for the survival of H. pylori when the external pH is nearly neutral (38). • • This finding leads us to speculate that the increased efficacy of quadruple therapy, in which bismuth and a proton pump inhibitor are given in combination with two antibiotics (17), is likely to be linked to this F1F0-ATPase-dependent survival at a neutral pH.Despite these subtle differences in the mechanisms for shutting down ATP synthesis, the effects are likely to be the same. • Moreover, the structural and compositional changes that we observed in the H. pylori outer membrane are likely to have direct consequences on the survival of these bacteria in the gastric mucosa. Reduced LPS synthesis could affect the stability of the bacterial glycocalyx (46) as well as increase the exposure of surface antigens, thereby rendering H. pylori more susceptible to host defenses and/or hydrophilic antimicrobial agents (50) • .Interestingly, considerable strain-dependent variations in the ability to preserve ATP levels were observed following culturing of the H. pylori strains in the presence of MICCBS and protective iron. • H. pylori 60190 ATP levels were considerably lower than those observed under both normal and iron-limiting culture conditions, and one explanation might be an increased requirement for energy by this strain. H. pylori 60190 carries the complete cag PAI, a locus of about 37 kb containing up to 31 genes (12). • Several of these genes code for a type IV secretion system (41) that includes an ATP-regulated inner membrane pore (53). • Supporting this hypothesis is the observation of higher levels of ATP in the other two strains grown under the same conditions. These strains do not possess the cag PAI (Tx-30a) or carry only a partial cag PAI (SS1) (14, 15). • In summary, our results suggest that whereas the antimicrobial effects of bismuth and iron deprivation on H. pylori may be similar, their respective mechanisms of intracellular action would appear to be mediated by different pathways within the cell. REFERENCES 1.al-Aoukaty, A., V. D. Appanna, and H. Falter. 1992. Gallium toxicity and adaptation in Pseudomonas fluorescens. FEMS Microbiol. Lett. 71:265-272. 2.Alarcon, T., D. Domingo, and M. Lopez-Brea. 1999. Antibiotic resistance problems with Helicobacter pylori. Int. J. Antimicrob. Agents 12:19-26. [PubMed] [Full Text]. 3.Andersen, L. P., H. Colding, and J. E. Kristiansen. 2000. Potentiation of the action of metronidazole on Helicobacter pylori by omeprazole and bismuth subcitrate. Int. J. Antimicrob. Agents 14:231-234. [PubMed] [Full Text]. 4.Armstrong, J. A., S. H. Wee, C. S. Goodwin, and D. H. Wilson. 1987. Response of Campylobacter pyloridis to antibiotics, bismuth and an acid-reducing agent in vitro— an ultrastructural study. J. Med. Microbiol. 24:343-350. [PubMed]. 5.Atherton, J. C., P. Cao, R. M. Peek, Jr., M. K. Tummuru, M. J. Blaser, and T. L. Cover. 1995. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 270:17771-17777. [PubMed] [Free Full Text 6.Baer, W., H. Koopmann, and S. Wagner. 1993. Effects of substances inhibiting or uncoupling respiratory-chain phosphorylation of Helicobacter pylori. Zentbl. Bakteriol. 280:253-258. 7.Beil, W., C. Birkholz, S. Wagner, and K. F. Sewing. 1995. Bismuth subcitrate and omeprazole inhibit Helicobacter pylori F1-ATPase. Pharmacology 50:333-337. [PubMed]. 8.Bereswill, S., S. Greiner, A. H. van Vliet, B. Waidner, F. Fassbinder, E. Schiltz, J. G. Kusters, and M. Kist. 2000. Regulation of ferritin-mediated cytoplasmic iron storage by the ferric uptake regulator homolog (Fur) of Helicobacter pylori. J. Bacteriol. 182:5948-5953. [Free Full text in PMC]. 9.Bereswill, S., F. Lichte, T. Vey, F. Fassbinder, and M. Kist. 1998. Cloning and characterization of the fur gene from Helicobacter pylori. FEMS Microbiol. Lett. 159:193-200. [PubMed] [Full Text]. 10.Blaser, M. J. 1992. Helicobacter pylori: its role in disease. Clin. Infect. Dis. 15:386-391. [PubMed] 11.Bode, G., F. Mauch, and P. Malfertheiner. 1993. The coccoid forms of Helicobacter pylori. Criteria for their viability. Epidemiol. Infect. 111:483-490. [PubMed]. 12.Censini, S., C. Lange, Z. Xiang, J. E. Crabtree, P. Ghiara, M. Borodovsky, R. Rappuoli, and A. Covacci. 1996. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA 93:14648-14653. [Free Full text in PMC]. 13.Coghlan, J. G., D. Gilligan, H. Humphries, D. McKenna, C. Dooley, E. Sweeney, C. Keane, and C. O'Morain. 1987. Campylobacter pylori and recurrence of duodenal ulcers—a 12-month follow-up study. Lancet 2:1109-1111. [PubMed]. 14.Cover, T. L., C. P. Dooley, and M. J. Blaser. 1990. Characterization of and human serologic response to proteins in Helicobacter pylori broth culture supernatants with vacuolizing cytotoxin activity. Infect. Immun. 58:603-610. [Free Full text in PMC]. 15.Crabtree, J. E., R. L. Ferrero, and J. G. Kusters. 2002. The mouse colonizing Helicobacter pylori strain SS1 may lack a functional cag pathogenicity island. Helicobacter 7:139-140. [PubMed] [Full Text]. 16.de Boer, W. A. 1999. Bismuth triple therapy: still a very important drug regimen for curing Helicobacter pylori infection. Eur. J. Gastroenterol. Hepatol. 11:697-700. [PubMed]. 17.de Boer, W. A., and G. N. Tytgat. 2000. Regular review: treatment of Helicobacter pylori infection. Br. Med. J. 320:31-34. [PubMed] [Free Full Text]. 18.Dhaenens, L., F. Szczebara, and M. O. Husson. 1997. Identification, characterization, and immunogenicity of the lactoferrin-binding protein from Helicobacter pylori. Infect. Immun. 65:514-518. [Free Full text in PMC]. 19.Domenico, P., L. Baldassarri, P. E. Schoch, K. Kaehler, M. Sasatsu, and B. A. Cunha. 2001. Activities of bismuth thiols against staphylococci and staphylococcal biofilms. Antimicrob. Agents Chemother. 45:1417-1421. [Free Full text in PMC] 20.Domenico, P., D. R. Landolphi, and B. A. Cunha. 1991. Reduction of capsular polysaccharide and potentiation of aminoglycoside inhibition in gram-negative bacteria by bismuth subsalicylate. J. Antimicrob. Chemother. 28:801-810. [PubMed]. 21.Domenico, P., J. Reich, W. Madonia, and B. A. Cunha. 1996. Resistance to bismuth among gram-negative bacteria is dependent upon iron and its uptake. J. Antimicrob. Chemother. 38:1031-1040. [PubMed]. 22.Domenico, P., J. M. Tomas, S. Merino, X. Rubires, and B. A. Cunha. 1999. Surface antigen exposure by bismuth dimercaprol suppression of Klebsiella pneumoniae capsular polysaccharide. Infect. Immun. 67:664-669. [Free Full text in PMC]. 23.Emery, T. 1986. Exchange of iron by gallium in siderophores. Biochemistry 25:4629-4633. [PubMed]. 24.Fekete, F. A., and L. L. Barton. 1991. Effects of iron(III) analogs on growth and pseudobactin synthesis in a chromiumtolerant Pseudomonas isolate. Biol. Met. 4:211-216. [PubMed]. 25.George, P. M., C. Conaghan, H. B. Angus, T. A. Walmsley, and B. A. Chapman. 1996. Comparison of histological and biochemical hepatic iron indexes in the diagnosis of genetic haemochromatosis. J. Clin. Pathol. 49:159-163. [PubMed]. • • • • • • • • • • • • • • • • • • • • • • • • • • • • 26.Goodwin, C. S., P. Blake, and E. D. Blincow. 1986. The minimum inhibitory and bactericidal concentrations of antibiotics and anti-ulcer agents against Campylobacter pylordis. J. Antimicrob. Chemother. 17:309-314. [PubMed]. 27.Goodwin, C. S., B. J. Marshall, E. D. Blincow, D. H. Wilson, S. Blackbourn, and M. Phillips. 1988. Prevention of nitroimidazole resistance in Campylobacter pylori by coadministration of colloidal bismuth subcitrate: clinical and in vitro studies. J. Clin. Pathol. 41:207-210. [PubMed]. 28.Huang, C. T., and P. S. Stewart. 1999. Reduction of polysaccharide production in Pseudomonas aeruginosa biofilms by bismuth dimercaprol (BisBAL) treatment. J. Antimicrob. Chemother. 44:601605. [PubMed] [Free Full Text]. 29.Keenan, J. I., and R. A. Allardyce. 2000. Iron influences the expression of Helicobacter pylori outer membrane vesicle-associated virulence factors. Eur. J. Gastroenterol. Hepatol. 12:1267-1273. [PubMed]. 30.Keenan, J. I., R. A. Allardyce, and P. F. Bagshaw. 1997. Dual silver staining to characterise Helicobacter spp. outer membrane components. J. Immunol. Methods 209:17-24. [PubMed] [Full Text]. 31.Koo, J., J. Ho, S. K. Lam, J. Wong, and G. B. Ong. 1982. Selective coating of gastric ulcer by tripotassium dicitrato bism uthate in the rat. Gastroenterology 82:864-870. [PubMed]. 32.Lambert, J. R., and P. Midolo. 1997. The actions of bismuth in the treatment of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 11(Suppl. 1):27-33. [PubMed]. 33.Lee, A., J. O'Rourke, M. C. De Ungria, B. Robertson, G. Daskalopoulos, and M. F. Dixon. 1997. A standardized mouse model of Helicobacter pylori infection: introducing the Sydney strain. Gastroenterology 112:1386-1397. [PubMed] [Full Text]. 34.Lee, F. I., I. M. Samloff, and M. Hardman. 1985. Comparison of tri-potassium di-citrato bismuthate tablets with ranitidine in healing and relapse of duodenal ulcers. Lancet 1:1299-1302. [PubMed]. 35.Leunk, R. D., P. T. Johnson, B. C. David, W. G. Kraft, and D. R. Morgan. 1988. Cytotoxic activity in broth-culture filtrates of Campylobacter pylori. J. Med. Microbiol. 26:93-99. [PubMed]. 36.Markwell, M. A., S. M. Haas, L. L. Bieber, and N. E. Tolbert. 1978. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 87:206-210. [PubMed]. 37.Marshall, B. J., J. A. Armstrong, G. J. Francis, N. T. Nokes, and S. H. Wee. 1987. Antibacterial action of bismuth in relation to Campylobacter pyloridis colonization and gastritis. Digestion 37:1630. [PubMed]. 38.McGowan, C. C., T. L. Cover, and M. J. Blaser. 1997. Analysis of F1F0-ATPase from Helicobacter pylori. Infect. Immun. 65:2640-2647. [Free Full text in PMC]. 39.McNulty, C. A., J. Dent, and R. Wise. 1985. Susceptibility of clinical isolates of Campylobacter pyloridis to 11 antimicrobial agents. Antimicrob. Agents Chemother. 28:837-838. [Free Full text in PMC]. 40.Midolo, P. D., J. R. Lambert, T. G. Kerr, and W. Tee. 1999. In vitro synergy between ranitidine bismuth citrate and tetracycline or clarithromycin against resistant strains of Helicobacter pylori. Eur. J. Clin. Microbiol. Infect. Dis. 18:832-834. [PubMed] [Full Text]. 41.Odenbreit, S., J. Puls, B. Sedlmaier, E. Gerland, W. Fischer, and R. Haas. 2000. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science 287:1497-1500. [PubMed] [Full Text]. 42.Ratledge, C., and L. G. Dover. 2000. Iron metabolism in pathogenic bacteria. Annu. Rev. Microbiol. 54:881-941. [PubMed] [Full Text] .43.Rauws, E. A., W. Langenberg, H. J. Houthoff, H. C. Zanen, and G. N. Tytgat. 1988. Campylobacter pyloridis-associated chronic active antral gastritis. A prospective study of its prevalence and the effects of antibacterial and antiulcer treatment. Gastroenterology 94:33-40. [PubMed]. 44.Sox, T. E., and C. A. Olson. 1989. Binding and killing of bacteria by bismuth subsalicylate. Antimicrob. Agents Chemother. 33:2075-2082. [Free Full text in PMC]. 45.Stoltenberg, M., M. Martiny, K. Sorensen, J. Rungby, and K. A. Krogfelt. 2001. Histochemical tracing of bismuth in Helicobacter pylori after in vitro exposure to bismuth citrate. Scand. J. Gastroenterol. 36:144-148. [PubMed] 46.Stratton, C. W. 1996. Mechanisms of action for antimicrobial agents: general principles and mechanisms for selected classes of antibiotics, p. 579-603. In V. Lorian (ed.), Antibiotics in laboratory medicine. Lippincott, Williams & Wilkins Co., Baltimore, Md. 47.Stratton, C. W., R. R. Warner, P. E. Coudron, and N. A. Lilly. 1999. Bismuth-mediated disruption of the glycocalyx-cell wall of Helicobacter pylori: ultrastructural evidence for a mechanism of action for bismuth salts. J. Antimicrob. Chemother. 43:659-666. [PubMed] [Free Full Text]. 48.Stryer, L. 1988. Biochemistry, 3rd ed. W. H. Freeman & Co., New York, N.Y 49.Tytgat, G. N. J., A. T. R. Axon, and M. F. Dixon. 1990. Helicobacter pylori: causal agent in peptic ulcer? Working Party Reports of the World Congress of Gastroenterology. Blackwell Scientific Publications Ltd., Oxford, England. 50.Vaara, M., and T. Vaara. 1983. Polycations sensitize enteric bacteria to antibiotics. Antimicrob. Agents Chemother. 24:107-113. [Free Full text in PMC]. 51.Van Caekenberghe, D. L., and J. Breyssens. 1987. In vitro synergistic activity between bismuth subcitrate and various antimicrobial agents against Campylobacter pyloridis (C. pylori). Antimicrob. Agents Chemother. 31:1429-1430. [Free Full text in PMC]. 52.Worst, D. J., B. R. Otto, and J. de Graaff. 1995. Iron-repressible outer membrane proteins of Helicobacter pylori involved in heme uptake. Infect. Immun. 63:4161-4165. [Free Full text in PMC]. 53.Yeo, H. J., S. N. Savvides, A. B. Herr, E. Lanka, and G. Waksman. 2000. Crystal structure of the hexameric traffic ATPase of the Helicobacter pylori type IV secretion system. Mol. Cell 6:1461-1472. Another YCP Controversy: Genotipificación of isolated clinical of Helicobacter pylori on the basis of associated genes to virulence . Correlation between Chilean types and others cultures(e.i.: Ethiopian) • Chilean Study: Genotipificación of isolated clinical of Helicobacter pylori on the basis of associated genes to virulence cagA, vacA and babA2. First isolation of a positive stock babA2. • Ethiopian study: Correlation of Prevalence between Helicobacter pylori vacA and cagA Genotypes in Others cultures. • Genotipificación of isolated clinical of Helicobacter pylori on the basis of associated genes to virulence cagA, vacA and babA2. First isolation of a positive stock babA2. • Apolinaria Garcia C1a, Ricardo Barra T1b, Thin Carolina Sch2, Fernando Kawaguchi P3, Natalia Trabal Fç, Sonia Montenegro H4d and Carlos González C1e 1 Laboratory of Bacterial Pathogenicity. Department of Microbiology, Faculty of Biological Sciences. 2 Department of Pathology and 3Departamento of Internal Medicine, Medicine Faculty. University of Conception. Conception, Chile. to 4Molecular Immunogenetics Laboratory, Ochsner Clinic Foundation, New Orleans, The USA. • Genotypying of clinical isolates of Helicobacter pylori by cagA, vacA and babA2 virulence associated genes. First isolation of a babA2 positive strain. • SUMMARY • Background. Gastroduodenal diseases associated to Helicobacter pylori species depends on host characteristics, environmental conditions and bacterial virulence factors, like cagA, vacA y babA2 genes product. Moreover, peptic ulcer disease is related with cagA+, vacAs1m1 strains while H. pylori cagA+, vacAs1 y babA2+ strains are associated to metaplasia and gastric cancer. Nonetheless, gene babA2 is still not described in clinical isolates from Chilean patients. • Aim. To investigate the presence of cagA, vacA (s and m) and babA2 genes in clinical isolates of H. pylori among Chilean patients. • Methods. Sixty six strains isolated from 41 patients were genotyped by PCR, using s1a, s1b, s2, m1, m2, cagA and babA2 primers described elsewhere. • Results. cagA gene was detected in 16 isolates (24.2%) while vacAs1a, vacAs1b, vacAs2, vacAm1, and vacAm2 were detected in 28 strains (42.4%), 14 strains (21.2%), 17 strains (25.8%), 21 strains (31.8%) and 29 strains (43.9%), respectively. • One strain (1.5%) was babA2 positive, being the first strain with this genotype described in Chile. This strain presented a genotype cagA+, vacAs1 y babA2+ which is related with metaplasia or gastric cancer. In addition, five strains showed an ulcerogenic profile cagA+, vacAs1m1. • Conclusion. The results presented indicate the prevalence of vacAs1m1 genotype among clinical strains analyzed, and the presence of low frequency of strains with babA2 + genotype. • Introduction. • The development of gastroduodenales pathologies associated to Helicobacter pylori has been associated to characteristics of the guest, the medio.ambiente and factors of virulence of the bacterium, such as codified by the genes cagA, vacA and babA2. • The ulcerous peptic disease is associated to stocks cagA+ and vacAs1m1, whereas the gastric presence of metaplasia and adenocarcinoma is related to stocks of H. pylori cagA+, vacAs1 and babA2+. • This last gene not yet has been described in Chilean stocks. • Objective. • Genotipificar isolated stocks of H. pylori of patients with gastroduodenal pathology, on the basis of the genes cagA, vacA (region s and m) and babA2. • Methods. • 66 stocks of H. were analyzed pylori obtained of gastric biopsies of 41 patients with high digestive pathology. • he genotipificación was made by means of PCR, using partidores for s1a, s1b, s2, m1, m2, cagA and babA2, described in Literature. • Results. Sixteen stocks presented/displayed the gene cagA (24.2%), 28 vacAs1a (42.4%), 14 vacAs1b (21.2%), 17 vacAs2 (25.8%), 21 vacAm1 (31.8%), 29 vacAm2 (43.9%) and one stock (1.5%) presented/displayed the gene babA2. Five stocks (7.6%) presented/displayed a genetic profile associate with ulcerogénicas stocks and the stock babA2+ (1.5%) presented/displayed a genotype associated to metaplasia and/or adenocarcinoma gastric. • • The infection by Helicobacter pylori establishes in the region antropìlorica of the stomach, in most of the infected individuals, a chronic, asintomática inflammation, long play, possibly because the immunological defensive mechanisms of the guest fail in their elimination (1, 2). • The persistence of the infection in certain patients is in the development of severe pathologies, that progress of chronic gastritis to peptic ulcer, atrófica gastritis, linfoma of gastric MALT and adenocarcinoma (3, 4, 5). • In spite of the high prevalence of the infection by H. pylori, only one minority of infected individuals develops a malignant severe pathology. This can be due to the genetic diversity between individuals (6); • • • to environmental factors, like type of diet, age of the first infection (7), and to specific factors of virulence of the bacterium (8,9). The gene cagA, that codifies a inmunodominante antigen, does not appear in all the stocks of H. pylori (10), but comprises of the pathogenicity small barren island (cagPAI), that contains 31 genes (11). • Therefore, its molecular detection indicates the presence of the PAI in the chromosome of the microorganism (9). • The stocks cag+ (stocks type I) are associated to greater virulence, when inducing visible gastric damage, whereas the stocks cag- (stocks type II) are associated to smaller virulence and they behave like bacteria at the table than more pathogenic (1). • • The vacuolizante citotoxina VacA is secretada by around 50% of the stocks of H. pylori and causes degeneration to vacuolar of the epithelial gastric cells and ulceración of the gastric mucosa (12). • The citotoxina is codified by the gene vacA, that can present/display genetic mosaic on the basis of alélicas variations in the regions mediates (alelos m1 or m2 and subtypes) and of signal (alelos s1 or s2 and subtypes) of the gene (13,14). • Specifically, one has demonstrated that stocks vacA s1/m1 have a high cytotoxic activity in comparison with stocks s1/m2 and that stocks s2/m2 would not have cytotoxic activity (13). • Recently one has demonstrated that factors of bacterial adhesion contribute to the pathogenicity of H. pylori (15-17). • Thus, the adhesina, codified by the gene babA2, favors a persistent union between the microorganism and the gastric epithelial cell by union of the bacterial cell through its BabA2 protein with group antigen Lewis B (LeB) present in the gastric mucosa (2). • Therefore, positive stocks of H. pylori babA2 present/display greater capacity of adhesion, however the negative stocks babA2 adhere weakly (18). • This adhesion has been associated with high levels of linfocitaria infiltration, intestinal glandular atrophy, metaplasia and increase of the epithelial proliferation, reporting a significant association with duodenal ulcer and gastric cancer (19). • According to Yu and col. (2002) (17), the gene babA2 could be a useful molecular marker to identify patients with greater risk of presenting/displaying associate severe pathologies to infections by H. pylori. • Alves Oliveira and col. (2003) (20) in a study in Brazil informed a strong association between babA2 and the presence into peptic ulcer or gastric carcinoma. • It is important to consider that the frequency of virulence determinants and its association with gastrointestinales pathologies vary considerably in different geographic regions.. • The presence of more than one of these genes of virulence would be associated with greater severity of the gastric injury. • One has inquired that stocks cagA+, vacAs1m1 would be associate of ulcers and stocks triple upon presentment positive cagA+, vacAs1, • babA2+ in addition to ulcer, would be associated with metaplasia and adenocarcinoma gastric (21, 22). • In Chile more of 90% of the patients with peptic ulcer and the 42 % of those who consult by dispepsia nonulcerosa they are infected with H. pylori (23). • In spite of the high prevalence of infection by H. pylori in our country, still the works in genotipificación of stocks including the genes are few cagA and vacA (24; 25; 26) and are not works with respect to the gene babA2. • The objective of the present study was to genotipificar isolated stocks of H. pylori of patients with gastroduodenal pathology, on the basis of the genes cagA, vacA (region s and m) including the gene babA2 not detected previously in Chilean stocks • Conclusion. • The results as a whole indicate the predominance of stocks with genotype vacAs1m1 between isolated clinical of H. pylori and a LF in the detection of the positive genotype babA2. • This gene could be explored as marking of more severe gastric injuries in our means. • • • • • • • • • • • • • • • • • • • • • • Referencias TAYLOR D, BLASER M J. The epidemiology of Helicobacter pylori infection. Epidemiol Rev 1991; 13: 42-59. NILSSON C, SILLEN A, ERIKSSON L, STRAND ML, ENROTH H, NORMARK S, FALK P, ENGSTRAND L. Correlation between cag pathogenicity island composition and Helicobacter pylori-associated gastroduodenal diseases. Infec Immun 2003; 71: 6573-81. MARSHALL B. Helicobacter pylori. Am J Gastroenterol. 1994; 89: 116-27. FOREMAN D, AND THE EUROGAST STUDY GROUP, An International Association between Helicobacter pylori infection and gastric cancer. Lancet 1993; 341: 359-62. WOTHERSPOON A, DOGLIONI C, DISS T, PAN L, MOSCHINI A, DE BONI M, ISAACSON P.. Regression of primary low-grade B-cell gastric lymphoma of mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori. Lancet.1993;342: 575-77. AZUMA T, ITO S, SATO F, YAMAZAKI Y, MIYAJI H, ITO Y, SUTO H, KURIYAMA M, KATO T, KHOLI Y. The role of the HLA-DQA1 gene in resistance to atrophic gastritis and gastric adenocarcinoma induced by Helicobacter pylori infection. Cancer 1998; 82: 1013-18. KATO I, VIVAS J, PLUMMER M, LOPEZ G, PERAZA S, CASTRO D, SANCHEZ V, CANO E, ANDRADE O, GARCIA R, FRANCESCHI S, OLIVER W, MUNOZ N. Environmental factors in Helicobacter pylori-related gastric precancerous lesions in Venezuela. Cancer Epidemiol Biomarkers Prev 2004; 13: 468-76. YANG J, WANG T, WANG H, KUO C, WANG J, WANG W. Genetic analysis of the cytotoxin associated gene and the vacuolating gene in Helicobacter pylori strains isolated from Taiwanese patients. Am J Gastroenterol 1997; 92: 1316-21. BLASER MJ, ATHERTON JC. Helicobacter pylori persistence: biology and disease. J Clin Invest 2004; 113: 321-33. COVACCI A, CENSINI S, BUGNOLI M. Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer. Proc Natl Acad Sci USA 1993; 90: 5791-95. 11. CENSINI S, LANGE C, XIANG Z, CRABTREE J, GHIARA P, BORODOVSKY M, RAPPUOLI R AND COVACCI A. Cag, a pathogenicity island of Helicobacter pylori, encoded type I-specific and disease-associated virulence factors. Proc Natl Acad Sci USA. 1996 ; 93: 14648-53. 12. COVACCI A, TELFORD JL, DEL GIUDICE G, PARSONNET J, RAPPUOLI R. Helicobacter pylori virulence and genetic geography. Science 1999; 284: 1328-33. 13. ATHERTON JC, CAO P, PEEK RM JR, TUMMURU MK, BLASER MJ, COVER TL. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J Biol Chem 1995; 270: 1771-7. 14. VAN DOORN LJ, FIGUEIREDO C, SANNA R, PENA AS, MIDOLO P, ATHERTON JC, BLASER MJ, QUINT W. Expanding allelic diversity of Helicobacter pylori vacA. J Clin Microbiol 1998; 36: 2597-2603. 15. MIZUSHIMA T, SUGIYAMA T, KOMATSU Y, ISHIZUKA J, KATO M, ASAKA M. Clinical relevance of the babA2 genotype of Helicobacter pylori in Japanese clinical isolates. J Clin Microbiol 2001; 39: 2463-65. 16. LAICH, KUO CH, CHEN YC, CHAO FY, POON SK, CJANG CS, WANG WC. High prevalence of cagA- and babA2-positive Helicobacter pylori clinical isolates in Taiwan. J Clin Microbiol 2002; 40:3860-62. 17. YU J, LEUNG WK, GO MYY, CJHAN MCW, TO KF, NG EKW, CHAN FKL, LING TKW, CHUNG SCS, SUNG JJY. Relationship between Helicobacter pylori babA2 status with gastric epithelial cell turnover and premalignant gastric lesions. Gut 2002; 51: 480-84. 18. BOREN T, NORMARK S, FALK P. Helicobacter pylori: molecular basis for host recognition and bacterial adherence. Trends Microbiol 1994; 2: 22128. 19. GERHARD M, LEHN N, NEUMAYER N. ET AL Clinical relevance of the Helicobacter pylori gene for blood-group antigen-binding adhesin. Proc Natl Acad Sci 1999; 96:1278-83. 20. ALVES O A, SANTOS A, BECATTINI G J, AGUIAR R G, CAMARGOS R A, OLIVEIRA C, DEMAS A, CABRAL M, FERREIRA N A y MAGALAHES Q D. babA2- and cagA-Positive Helicobacter pylori Strains Are Associated with Duodenal Ulcer and Gastric Carcinoma in Brazil. J Clin Microbiol 2003; 41: 3964–66. • • • • • • • • • • • • • • • • • • 21. GERHARD M, LEHN N, NEUMAYER N ET AL. Clinical relevance of the Helicobacter pylori gene for blood-group antigen-binding adhesin. Proc Natl Acad Sci USA. 1996; 96: 12778-83. 22. ZAMBON CF, NAVAGLIA F, BASSO D, RUGGE M, PLEBANI M. Helicobacter pylori babA2, cagA, and s1vacA genes work synergistically in causing intestinal metaplasia. J Clin Pathol 2003; 56: 287-91. 23. PRADO, V. Enfermedades infecciosas emergentes: ¿Un problema nuevo? Rev Med Chile 1996; 124: 7-10. 24. MARTÍNEZ A, GONZÁLEZ C, KAWAGUCHI F, MONTOYA R, CORVALÁN A, MADARIAGA J, ROA J, GARCÍA A, SALGADO F, SOLAR H, PALMA M. Helicobacter pylori: análisis de cagA y genotipificación de vacA en Chile. Detección de una cepa s2/m1. Rev Med Chile 2001; 129: 1147-53. 25. FAÚNDEZ G, TRONCOSO M, FIGUEROA G. cagA and vacA in strains of Helicobacter pylori from ulcer and non-ulcerative dyspepsia patients. BMC Gastroenterol 2002, 2: 20. 26. ARAYA J, ANABALÓN L, ROA I, BRAVO M, VILLASECA M, GUZMÁN P, ROA J. Relación de la genotipificación de Helicobacter pylori con la forma e intensidad de la gastritis en población adulta portadora de patología gástrica benigna. Rev Med Chile 2004; 132: 1345-54. 27. WORKING PARTY OF THE EUROPEAN HELICOBACTER PYLORI STUDY GROUP. Guidelines for clinical trials in Helicobacter pylori infection. Gut 1997; 41 (Suppl 2): S3. 28. MAZURIER S, VAN DE GIESSEN A, HEUVELMAN K, WERNARS K. RAPD analysis of Campylobacter isolates: DNA fingerprinting without the need to purify DNA. Lett Appl Microbiol 1992; 14: 260-62. 29. TUMMURU MK, COVER TL, BLASER MJ. Cloning and expression of a high-molecular-mass major antigen of Helicobacter pylori: evidence of linkage to cytotoxin production. Infect Immun 1993; 61: 1799-809. 30. SALGADO F, GARCÍA A, OÑATE A, GONZÁLEZ C, KAWAGUCHI F. Increased in-vitro and in-vivo biological activity of lipopolysaccharide extracted from clinical low virulence vacA genotype Helicobacter pylori strains. J Med Microbiol 2002; 51: 771-76. 31. NAVAGLIA F, BASSO B y PLEBANI M. Touchdown PCR: a rapid method to genotype Helicobacter pylori infection. Clin Chim Acta 1997; 262: 57-60. 32. PODZORSKI R, PODZORSKI D, WUERTH A, TOLIA V. Analysis of the vacA, cagA, cagE, iceA and babA2 genes in Helicobacter pylori from sixty-one pediatric patient from the Midwestern united states. Diagnos Microbiol Infec Dis. 2003, 46: 83-88. 33. HEHENBERGER P, GRETECHEL S. Gastric Cancer. The Lancet, 2003. Tomo 362, N 9380: pp 305. 34. KIM S-Y, WOO C, LEE Y-M, SON B, KIM J, CHAE H, YOUN S, PARK S. Genotyping cagA, vacA subtype, iceA and babA of Helicobacter pylori isolates from Korean patients, and their association with gastroduodenal diseases. J Korean Med Sci 2001; 16: 579-84. 35. LAI C-H, KUO C-H, CHEN Y-C, CHAO F-Y, POON S-K, CHANG C-S AND WANG W-C. High prevalence of cagA and babA2 positive Helicobacter pylori clinical isolates in Taiwan. J Chin Med Assoc 2002; 40: 3860-62. 36. YAKOOB J, FAN X G, PENG X N, HU G L, ZHANG Z. Helicobacter pylori cagA and vacA cytotoxin genes in Changsha, China. Br J Biomed Sci 2002; 59: 150-3. 37. YU J, LEUNG WK, GO MYY, CHAN MCW, TO KF, NG EKW, CHAN FKL, LING TKW, CHUNG SCS, SUNG JJK. Relationship between Helicobacter pylori babA2 status with gastric epithelial cell turnover and premalignant gastric lesions. Gut. 2002; 51: 480-484. Correlation of Prevalence between Helicobacter pylori vacA and cagA Genotypes in Others cultures (e.i. : Ethiopian) DRA. ALEJANDRA MARTINEZ, IGNACIO ALFARO, TAMARA PEREZ, CINTHYA PEREZ. • A total of 300 gastric biopsy samples and 50 Helicobacter pylori isolates were collected from Ethiopian adult dyspeptic patients. • The vacA and cagA genes were detected in 90 and 79% of biopsy specimens, • respectively, and in 100 and 87% of clinical isolates, respectively. Both genes were detected in 84% of the gastric biopsy samples and in 87% of the clinical isolates. • Among vacA genotypes, the s1/m1 genotype was the most common in gastric biopsy samples (48%). • The vacA and cagA positive H. pylori strains were • detected to a higher degree in patients with chronic active gastritis (71%) than patients with other histopathological findings (29%) (P < 0.05). • • • • • • • Several Helicobacter pylori virulence genes related to the risk of gastroduodenal diseases have been proposed. The vacuolating cytotoxin (vacA) gene is present in virtually all H. pylori strains and contains at least two variable regions, the signal (s) region, which encodes the signal peptide, and the middle (m) region (4). • • • The s region has been divided into two subtypes, s1 and s2, and the m region has been divided into two subtypes, m1 and m2 (19). • • • • • • • • The amount of cytotoxin produced is highest with the s1/m1 allele, followed by the s1/m2 allele, while no cytotoxin activity is found when s2/m2 is present (19). The cytotoxin-associated gene (cagA) is a marker for a genomic pathogenicity island of 40 kb (6). A significant association between the presence of ulcers or gastric carcinoma and the presence of vacA type s1 and cagA gene (5, 19). • • • • The present study represents the first in Ethiopia to detect H. pylori vacA and cagA genotypes from gastric biopsy samples and clinical isolates using PCR-based methods. MATERIALS AND METHODS • • • • • • • • • • • • Study subjects. A total of 300 consecutive informed and consenting adult patients with dyspeptic symptoms from the gastrointestinal referral and followup clinics of Department of Internal Medicine, Tikur Anbassa University Hospital, Addis Ababa, Ethiopia, were investigated for H. pylori between November 2000 and August 2002. The mean age of the patients was 36.5 years (standard deviation, 13.8 years; range, 15 to 90 years). The majority of patients (76%) were between the ages of 15 and 44 years. Of the 300 patients, 186 (62%) were males and 114 (38%) were females (resulting in an overall male to female ratio of 1.6:1). The study was approved by the Department Graduate Committee, the Faculty Research Publications Committee and endorsed by the Faculty Academic Commission and has been ethically cleared. Culture and identification • • • • • • • • • • • • • • • • • • • • • . Antral gastric biopsy samples were taken from each dyspeptic patient. The biopsy specimens were put into sterile phosphatebuffered saline containing 15% glycerol and immediately transported to laboratory for culture. Biopsy samples for molecular analysis were kept frozen in 15% tryptone soy broth (Oxoid Ltd., Basingstoke, England) and stored at _70°C until analyzed. H. pylori was cultured from antral biopsy specimens using a standard method (17). H. pylori identification was based on morphology, Gram staining, oxidase, catalase, and urease tests. All the isolated H. pylori strains were kept frozen at _70°C in the tryptone soy broth medium containing 15% (vol/vol) glycerol until genotyping was performed. The H. pylori reference strain (CCUG 17874) (Culture Collection, University of Gothenburg, Gothenburg, Sweden) was cultured throughout the study for quality control. Histopathology. Gastric biopsy specimens were fixed in 10% formalin and embedded in paraffin. The sections (4 to 5 _m thick) were cut and stained with hematoxylin and eosin (2). The histological findings from the sections stained with hematoxylin and eosin were scored according to the updated Sydney system of classification and grading of gastritis (7). Genomic DNA extraction. Biopsy specimens and isolates were centrifuged at • • • • • • • • 10,000 _ g for 5 min. The DNA was extracted from the pellets by use of the QIAamp DNA kit (QIAGEN, Hilden, Germany) according to the manufacturer’s recommendations and DNA stored at _20°C until analysis. DNA extraction negative controls were performed in parallel by including sterile tubes without samples to check for contamination of the DNA extraction reagents. PCR-DGGE. The PCR amplification was carried out using a GeneAmp 2700 Thermal cycler (Applied Biosystems, Foster City, Calif.). • • • • • • A seminested Helicobacter genus-specific PCR assay targeting the 16S rDNA was used to amplify Helicobacter DNA (9). Denaturing gradient gel electrophoresis (DGGE) analysis of the PCR products was performed in a DCode system (Bio-Rad, Hercules, Calif.) as recently described (1). Migration ladder containing PCR products of reference Helicobacter strains (H. muridarum [CCUG 29262], H. bilis [CCUG 38995], H. pullorum [NCTC 12825], H. pylori [CCUG 17874], “Flexispira rappini” [CCUG 23435], H. hepaticus [CCUG 33637], and H. bizzozeronii [AF 53]) was run in parallel as a mobility ladder. vacA and cagA genotyping. • • • • • • • • Detection of H. pylori vacA and cagA genes was performed on gastric biopsy specimens and isolates positive for H. pylori by PCR-DGGE as previously described (15, 19). As a positive control, H. Pylori (CCUG 17874) DNA (_0.1 ng) was added to the reaction mixture, while 5 _l of sterile deionized Millipore-filtered water was added to the reaction mixture as a negative control. Estimation of size of the PCR products was done by using Gene ruler 100-bp DNA ladder (Fermentas, Vilnius, Lithuania). The products of each PCR assay were visualized by electrophoresis in a 1.5% agarose gel containing ethidium bromide (15). RESULTS AND DISCUSSION • • • Histopathological examinations were performed on 276 (92%) of the gastric biopsy specimens, whereas the remaining 24 (8%) were not adequate (too small). • Abnormal findings were observed in all examined specimens. • • Chronic gastritis was found in 48 (17.4%) patients, chronic active gastritis in 185 (67%), chronic atrophic gastritis in 24 (9%); • • chronic atrophic gastritis with intestinal metaplasia in 17 (6%) and malignant lesions in 2 (0.6%) patients. • • The overall prevalence of chronic gastritis was 99.3%. The most common histopathological findings in the present study were similar to those reported from other parts of Africa (10). • • • figures reported from The Netherlands (36%), Hong Kong (26 to 31%), and Nigeria (24%), but lower than figures reported from Brazil (80%) and Korea (78%) (3, 11, 16, 20, 21). In the • The seminested Helicobacter genus-specific PCR assay detected Helicobacter DNA in 273 of 300 (91%) of the biopsy specimens. DGGE analysis showed that all PCR products have • mobility pattern similar to the H. pylori reference strain (Fig. 1). • The vacA gene was detected in 246 of 273 (90%) of H. pylori-positive gastric biopsy specimens, which is similar to reported results from The Netherlands (93%) and Hong Kong • (95.8%) (14, 21), emphasizing high sensitivity of the PCR method employed in the present study. • The vacA genotypes s1/m1, s1/m2, s2/m2, and s2/m1 were found in 48, 28, 9, and 2% of the specimens, respectively (Table 1), whereas, 15 biopsy specimens (6%) were incomplete and thus did not yield a detectable PCR product for the vacA s or m regions. • The pattern of vacA alleles in this study is in agreement with those reported in other studies (3, 8, 16, 18, 20). • However, the frequencyof vacA s1/m1 allelic type in this study is higher than • • • In the present investigation, the rare vacA s2/m1 allele was detected in 4 (2%) of the 246 gastric biopsy specimens examined, also reported in studies in South Africa and Chile (12, 13). • • • Multiple vacA genotypes were found in 18 (7%) of the 246-biopsy specimens examined. The most frequent multiple vacA genotypes were s1/m1m2 (11 of 18; 61%). The vacA was detected in all 52 H. pylori isolates tested (Table 1). • The prevalence of the vacA subtypes s1/m1, s1/m2, and s2/m2 was 60, 27, and 7%, respectively. Three (6%) of the isolates contained mixed vacA subtypes; two s1/m1m2 and one s1s2/m2 from a single H. pylori isolate. The multiple vacA genotypes detected in this study are similar to results from Italy reported by Blaser and Berg (5). Surprisingly, the prevalence of multiple vacA genotypes in this study was much lower compared with results reported from Brazil (13%), Chile (32%), Korea (18%), and The Netherlands (11%) (3, 11, 13, 20). • • • • • • The low prevalence of multiple vacA genotypes in a country with a high prevalence of H. pylori infections in the general population may be the result of a low number of mosaics of any combination of signal (s) and mid-region (m) alleles of the bacteria circulating in the community. • Of the 273 H. pylori PCR-positive biopsy specimens, 217 • (79%) were cagA positive. • Four different genotypic combinations were recognized based on analysis of the positive and negative vacA and cagA results—vacA_ cagA_, vacA_ cagA, vacA cagA, and vacA cagA_, which were found in 76, 14, 6, and 4% of specimens, respectively (Table 2). • Forty-five of the 52 (87%) H. pylori strains were positive for both vacA and cagA, whereas the remaining isolates 7 (13%) were only vacA positive(Table 2). • Statistical analysis showed no difference in the detection of the vacA and cagA in gastric biopsy specimens and clinical isolates (P 0.70 for vacA, P 0.96 for cagA). • In • addition, no statistical differences in the frequency of detection • of the different vacA allelic types from gastric biopsy specimens • and clinical isolates were found (P _ 0.05). • The prevalence of • The prevalence of cagA positive H. pylori strains varies from one geographic region • to another, e.g., 38% in Chile, 48% in Sri Lanka, 67% in The Netherlands, 81% in the United Sates, 90% in Hong Kong, 97% in Korea, 93% in Nigeria and 94% in Brazil (3, 8, 11, 13, 16, 18, 20, 21). • Correlation of histopathology results with vacA and cagA genotypes showed that vacA and cagA positive strains were detected to a higher degree in patients with chronic active gastritis (71%) compared with patients with • other histopathological findings (29%) (P _ 0.05) (Table 3). • • • • Molecular analyses demonstrated that more than 80% of the FIG. 1. DGGE analysis of PCR products amplified by using Helicobacter genus-specific primers. Lanes, 1 to 4 and 6 to 22, H. Pyloripositive samples; 5, H. pylori-negative sample; M, mobility marker • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • REFERENCES 1. Abu Al-Soud, W., M. Bennedsen, S. L. W. On, I.-S. Ouis, P. Vandamme, H.-O. Nilsson, A. Ljungh, and T. Wadstro¨m. 2003. Assessment of PCRDGGE for the identification of diverse Helicobacter species, and application to faecal samples from zoo animals to determine helicobacter prevalence. J. Med. Microbiol. 52:765–771. 2. Anim, J. T., N. Al-Sobkie, A. Prasad, B. John, P. N. Sharma, and I. AlHamar. 2000. Assessment of different methods for staining Helicobacter pylori in endoscopic gastric biopsies. Acta Histochem. 102:129–137. 3. Ashour, A. A., P. P. Magalhaes, E. N. Mendes, G. B. Collares, V. R. de Gusmao, D. M. Queiroz, A. M. Nogueira, G. A. Rocha, and C. A. de Oliveira. 2002. Distribution of vacA genotypes in Helicobacter pylori strains isolated from Brazilian adult patients with gastritis, duodenal ulcer or gastric carcinoma. FEMS Immunol. Med. Microbiol. 33:173–178. 4. Atherton, J. C., P. Cao, R. M. Peek, Jr., M. K. R. Tummuru, M. J. Blaser, and T. L. Cover. 1995. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. J. Biol. Chem. 270:17771–17777. 5. Blaser, M. J., and D. E. Berg. 2001. Helicobacter pylori genetic diversity and risk of human disease. J. Clin. Investig. 107:767–773. 6. Censini, S., C. Lange, Z. Xiang, J. E. Crabtree, P. Ghiara, M. Borodovsky, R. Rappuoli, and A. Covacci. 1996. Cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA 93:14648–14653. 7. Dixon, M. F., R. M. Genta, J. H. Yardley, and P. Correa. 1996. Classification and grading of gastritis. The updated Sydney system. International Workshop on the Histopathology of Gastritis, Houston 1994. Am. J. Surg Pathol. 20:1161–1181. 8. Fernando, N., J. Holton, D. Vaira, M. DeSilva, and D. Fernando. 2002. Prevalence of Helicobacter pylori in Sri Lanka as determined by PCR. J. Clin. Microbiol. 40:2675–2676. 9. Goto, K., H. Ohashi, A. Takakura, and T. Itoh. 2000. Current status of helicobacter contamination of laboratory mice, rats, gerbils, and house musk shrews in Japan. Curr. Microbiol. 41:161–166. 10. Kidd, M., J. A. Louw, and I. N. Marks. 1999. Helicobacter pylori in Africa: observations on an enigma within an enigma’. J. Gastroenterol. Hepatol. 14:851–858. 11. Kim, S. Y., C. W. Woo, Y. M. Lee, B. R. Son, J. W. Kim, H. B. Chae, S. J. Youn, and S. M. Park. 2001. Genotyping cagA, vacA subtype, iceA1, and babA of Helicobacter pylori isolates from Korean patients, and their association with gastroduodenal diseases. J. Korean Med Sci. 16:579–584. 12. Letley, D. P., A. Lastovica, J. A. Louw, C. J. Hawkey, and J. C. Atherton. 1999. Allelic diversity of the Helicobacter pylori vacuolating cytotoxin gene in South Africa: rarity of the vacA s1a genotype and natural occurrence of an s2/m1 allele. J. Clin. Microbiol. 37:1203–1205. • • • • 13. Martinez, A., C. Gonzalez, F. Kawaguchi, R. Montoya, A. Corvalan, J. Madariaga, J. Roa, A. Garcia, F. Salgado, H. Solar, and M. Palma. 2001. Helicobacter pylori: cagA analysis and vacA genotyping in Chile. Detection of a s2/m1 strain. Rev. Med. Chil. 129:1147–1153. • • • • 14. Scholte, G. H., L. J. van Doorn, W. G. Quint, and J. Linderman. 2001. Genotyping of Helicobacter pylori strains in formalin-fixed or formaldehydesublimatefixed paraffin-embedded gastric biopsy specimens. Diagn. Mol. Pathol. 10:166–170. • • • • • • • • • • • • • • • • • • • • • • • • • • 15. Sjunnesson, H., T. Falt, E. Sturegard, W. Abu Al-Soud, A. Ljungh, and T. Wadstro¨m. 2003. PCR-denaturing gradient gel electrophoresis and two feces antigen tests for detection of Helicobacter pylori in mice. Curr. Microbiol. 47:278–285. 16. Smith, S. I., C. Kirsch, K. S. Oyedeji, A. O. Arigbabu, A. O. Coker, E. Bayerdoffer, and S. Miehlke. 2002. Prevalence of Helicobacter pylori vacA, cagA and iceA genotypes in Nigerian patients with duodenal ulcer disease. J. Med. Microbiol. 51:851–854. 17. Soltesz, V., B. Zeeberg, and T. Wadstro¨m. 1992. Optimal survival of Helicobacter pylori under various transport conditions. J. Clin. Microbiol. 30:1453– 1456. 18. Straus, E. W., H. Patel, J. Chang, R. M. Gupta, V. Sottile, J. Scirica, G. Tarabay, S. Iyer, S. Samuel, and R. D. Raffaniello. 2002. H. pylori infection and genotyping in patients undergoing upper endoscopy at inner city hospitals. Dig. Dis. Sci. 47:1575–1581. 19. van Doorn, L. J., C. Figueiredo, R. Rossau, G. Jannes, M. van Asbroeck, J. C. Sousa, F. Carneiro, and W. G. V. Quint. 1998. Typing of Helicobacter pylori vacA gene and detection of cagA gene by PCR and reverse hybridization. J. Clin. Microbiol. 36:1271–1276. 20. van Doorn, L. J., C. Figueiredo, R. Sanna, A. Plaisier, P. Schneeberger, W. de Boer, and W. Quint. 1998. Clinical relevance of the cagA, vacA, and iceA status of Helicobacter pylori. Gastroenterology 115:58–66. 21. Wong, B. C., Y. Yin, D. E. Berg, H. H. Xia, J. Z. Zhang, W. H. Wang, W. M. Wong, X. R. Huang, V. S. Tang, and S. K. Lam. 2001. Distribution of distinct vacA, cagA and iceA alleles in Helicobacter pylori in Hong Kong. Helicobacter 6:317–324. Departamento de Microbiología Facultad de Ciencias Biológicas Universidad de Concepción CITOMETRÍA DE FLUJO EN LA EVALUACIÓN DE POTENCIAL DE MEMBRANA Y VIABILIDAD CELULAR DE Helicobacter pylori. T.M. Juan Luis Castillo Navarrete, Dra Apolinaria García