4.1 – Capturing Solar Energy: The Light
advertisement

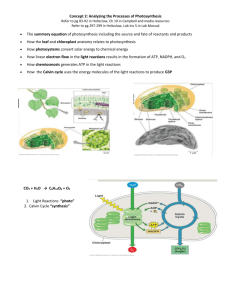
4.1 – Capturing Solar Energy: The Light-Dependent Reactions (Taken from Biology 12, MHR, 2011) Photosynthesis is carried out by plants, algae, photosynthetic protists, and cyanobacteria to produce their own food. The energy from the sun needs to be converted into a form of chemical energy that can be used by all non-photosynthetic organisms. The most common chemical energy produced from photosynthesis is glucose. If you recall, glucose is also the most common molecule that organisms use for fuel in the process of cell respiration. The overall equation for photosynthesis is: 6CO2 + 6H2O + light energy C6H12 O6 + 6O2 2 Stages of Photosynthesis Photosynthesis takes place within the chloroplasts. Stage 1: Light-Dependent Reactions. Capturing light energy and using that captured light energy to synthesize ATP and reduced NADP+ (NADPH). Stage 2: Light-Independent Reactions (The Calvin cycle or carbon fixation). Using the free energy of ATP and the reducing power of NADPH to synthesize organic compounds, such as glucose, from CO2 and H2O. Stage 1: Light-Dependent Reactions Some of the light energy absorbed by chlorophyll is used to produce ATP. Some of the light energy absorbed by chlorophyll is used to split water molecules. This is called photolysis of water. Photolysis of water results in the formation of oxygen and hydrogen. The oxygen is released as a waste product. (Damon, McGonegal, Tosto, Ward, 2007, pg 74-75), (Carter-Edwards, Gerards, Gibbons, McCallum, Noble, Parrington, Ramlochan, Ramlochan, 2011), (Allott, 2007, pg 21) Stage 2: Light-Independent Reactions Carbon dioxide is absorbed for use in photosynthesis. The carbon from the carbon dioxide is used to make a wide range of useful organic molecules for the plant (e.g., glucose). The conversion of carbon in a gas to carbon in solid compounds is called carbon fixation. Carbon fixation involves the use of hydrogen from photolysis and energy from ATP. Chloroplasts Chloroplasts have an outer membrane and an inner membrane. The interior space contains a semiliquid material called stroma with a system of membrane-bound sacs called thylakoids, some of which are stacked on top of one another to form grana. Thylakoid membranes contain chlorophyll molecules and electron transport chains. See Figure 4.2, pg 157. Nature of Light Light is a form of energy that travels at 3 x 108 m/s in the form of wave packets called photons. Photons are characterized by a wavelength that is inversely proportional to their energy. Photons with short wavelengths have high energy and those with long wavelengths have low energy. Different wavelengths appear as different colours. (e.g.) A wavelength of 400-525nm (violet-blue light), 525-625 (green-yellow light), and 625-700nm (orange-red light) form the visible part of the spectrum. Plants contain a variety of pigments in chloroplasts. Chlorophyll is the photosynthetic pigment that dominates in most plant species. For the purpose of considering how chlorophyll absorbs light, we will consider the blue end spectrum, the green middle spectrum, and the red end spectrum. Chlorophyll, the green pigment common to all photosynthetic cells, absorbs all wavelengths of visible light, except green, which it reflects. When sunlight hits a leaf, the red and blue wavelengths of light are absorbed by chlorophyll and used for photosynthesis. Almost all the energy of the green wavelengths is reflected not absorbed. This is why the plant appears green. Chlorophyll contains a porphyrin ring with a magnesium atom in the middle. Chlorophyll also has a long hydrocarbon tail (hydrophobic) that anchors the molecule into a membrane. Chlorophyll a is the only pigment that can transfer the energy of light to the carbon fixation reactions of photosynthesis. Chlorophyll b and the carotenoids act as accessory pigments, transferring their energy to chlorophyll a. (Damon, McGonegal, Tosto, Ward, 2007, pg 74-75), (Carter-Edwards, Gerards, Gibbons, McCallum, Noble, Parrington, Ramlochan, Ramlochan, 2011) Action Spectrum and Absorption Spectrum of Photosynthesis Biology 12 Blackline Masters BLM 4-8 Why Chlorophyll Appears Green Copyright © 2011 McGraw-Hill Ryerson Limited 978-0-07-106045-5 The diagram above shows the absorption spectra of some typical photosynthetic pigments. It shows the relative amounts of light absorbed at the different wavelengths of visible light. The energy that drives photosynthesis comes from light therefore the wavelength of the light absorbed by the chloroplasts partly determines the rate of photosynthesis. Action spectrum is the rate of photosynthesis at particular wavelengths of visible light. To produce an action spectrum for a particular plant, the rate of photosynthesis is determined by measuring oxygen production. The higher the oxygen production, the higher the rate of photosynthesis. (Damon, McGonegal, Tosto, Ward, 2007, pg 234) (Burrell, J. G. (2002-11) Click4Biology (version 0820.2011). Thailand: Bangkok; URL http://click4biology.info) By looking at the 2 graphs above (absorption spectrum and action spectrum), you can see two correlations: 1) Blue light and red light show the greatest absorption and they also represent the peaks in the rate of photosynthesis. 2) Green light shows low absorption and a lower rate of photosynthesis. (Remember: the green light is reflected. That is why the plants appear green.) (Damon, McGonegal, Tosto, Ward, 2007, pg 234) There are some differences between the action spectrum and the absorption spectra. If you look at the green-yellow range of both graphs, you can see that little light is absorbed but there is some photosynthesis. This is due to accessory pigments, including xanthophylls and carotene, which absorb wavelengths that chlorophyll cannot. (Allott, 2007, pg 77) Stomata Carbon dioxide and water that are used to synthesize glucose through photosynthesis are taken up by the leaf and then enter into plant cells and chloroplasts. Water enters the leaf through veins, and carbon dioxide enters via openings called stomata. Stomata are open in the daytime and closed at night. Light-activated proton pumps in guard cell membranes cause K+ ions to move from neighbouring epidermal cells into guard cells. As a result, water moves by osmosis into guard cells, causing them to swell. Increasing turgor pressure within guard cells causes their membranes to buckle and stomata to open. As the concentration of sucrose in guard cells decreases in the evening, water moves out of the cells and stomata close. See Figure 4.2, pg 157 Overview of Photosynthesis The overall equation for photosynthesis is: 6CO2 + 6H2O + light energy C6H12 O6 + 6O2 Light-Dependent Reactions occurs in the thylakoid membranes of the chloroplast Clusters of photosynthetic pigments, called photosystems, (found in thylakoid membrane) absorb photons of particular wavelengths and through the light reactions, transfer their energy to ADP and NADP+, forming ATP and NADPH. Photosystem II absorbs light with a wavelength of 680nm. Photosystem I absorbs light with a wavelength of 700nm. 1) 2) The light-dependent reactions begin when photons strike a photosynthetic membrane. This process may be divided into 3 parts: Photoexcitation: absorption of a photon by an electron of chlorophyll Electron Transport: transfer of the excited electron through a series of membranebound electron carriers, resulting in the pumping of a proton through the photosynthetic membrane, which creates an H+ reservoir and eventually reduces an electron acceptor 3) Chemiosmosis: the movement of protons through ATPase complexes to drive the phosphorylation of ADP to ATP Non-cyclic Electron Flow and Chemiosmosis See Figure 4.7, pg 160 and Figure 4.8, pg 161 In the light-dependent reactions, photosystem II passes electrons to photosystem I via an electron transport system, which contains the b6-f complex. This complex acts as a proton pump to produce a proton gradient across the thylakoid membrane. The electrons lost from the reaction centre of photosystem II are replenished by the oxidation of water. Photosystem I uses the electrons to reduce NADP+ to NADPH. The passage of one electron pair through ETC generates 1 NADPH and slightly more than 1 ATP. This ratio is not enough for the light-independent reactions. They require 3 ATP molecules to 2 NADPH molecules. Step 1 The P680 molecule in the reaction centre of photosystem II absorbs a light photon, exciting an electron. When the excited electron leaves P680 in photosystem II and goes to the electron acceptor, P680 is missing an electron. A water-splitting enzyme takes two water molecules and removes four electrons, four H+, and 2 oxygen atoms. This process is known as photolysis. P680+ accepts these electrons one at a time to replace the missing electrons. P680+ then absorbs another photon, becomes reduced, and passes on another electron. This process occurs four times to produce one O2 molecule. The four H+ from the two water molecules remain in the thylakoid space. The O atoms from the water molecules immediately form an O2 molecule. This is the oxygen that plants release into the environment as a waste product. Step 2 From the electron acceptor, the energized electrons are transferred, one at a time, along a series of electron-carrying molecules known as an electron transport system. With each transfer of electrons along the ETC, a small amount of energy is released. The released energy is used by b6-f complex to pump H+ from the stroma across the thylakoid membrane, into the thylakoid lumen. Eventually, there is an accumulation of H+ in the thylakoid lumen. This generates an H+ concentration gradient across the thylakoid membrane. Step 3 At the same time as steps 1 and 2 are taking place, light energy is absorbed by photosystem I. This energy travels to the reaction centre P700 molecule, where electrons become excited. Again, the excited electrons are passed to a high-energy electron acceptor. In photosystem I, the lost electrons are replaced by those that have reached the end of the electron transport system from photosystem II. Step 4 The electrons that were received by the electron acceptor from photosystem I are used by the enzyme NADP reductase to reduce NADP+ to form NADPH. NADPH will be used in the light-independent reactions (Calvin Cycle). See Figure 4.8, pg 161 Making ATP by Chemiosmosis ATP synthesis in the light-dependent reactions uses the same mechanism as ATP synthesis in aerobic respiration – chemiosmosis (movement of hydrogen ions). Photophosphorylation: the use of photons of light to drive the phosphorylation of ADP to produce ATP via chemiosmosis H+ ions that are pumped from the stroma to the thylakoid space can’t diffuse back across membrane because the membrane is impermeable to H+ ions. The chloroplast ATP synthase provides the only pathway for the H+ to move down the concentration gradient. This pathway is linked to a mechanism that bonds a free phosphate group to an ADP molecule to form ATP. As the H+ move down their concentration gradient through ATP synthase, the energy of the gradient is used to produce ATP molecules. (Carter-Edwards, Gerards, Gibbons, McCallum, Noble, Parrington, Ramlochan, Ramlochan, 2011) NADPH and ATP are the final products of the light-dependent reaction. They supply the chemical energy needed for the light-independent reaction to occur. Since light is needed for the production of ATP in photosynthesis, it is referred to as photophosphorylation. Comparison of Chemiosmosis in Cellular Respiration and Photosynthesis Respiration Chemiosmosis 1) involves an electron transport chain embedded in the membranes of the cristae 2) Energy is released when electrons are exchanged from one carrier to another. 3) Released energy is used to actively pump hydrogen ions into the intermembrane space. 4) Hydrogen ions come from the matrix. 5) Hydrogen ions diffuse back into the matrix through the channels of ATP synthase. 6) ATP synthase catalyzes the oxidative phosphorylation of ADP to form ATP. Photosynthesis Chemiosmosis 1) involves an electron transport chain embedded in the membranes of the thylakoids 2) Energy is released when electrons are exchanged from one carrier to another. 3) Released energy is used to actively pump hydrogen ions into the thylakoid space (lumen). 4) Hydrogen ions come from the stroma. 5) Hydrogen ions diffuse back into the stroma through the channels of ATP synthase. 6) ATP synthase catalyzes the photophosphorylation of ADP to form ATP. (Damon, McGonegal, Tosto, Ward, 2007, pg 230) Cyclic Electron Flow When light is not the limiting factor, NADPH tends to accumulate in the stroma and there is a shortage of NADP+. The normal flow of electrons in the thylakoid membrane is inhibited because NADP+ is needed as a final acceptor of electrons. Cyclic photophosphorylation allows ATP production when NADP+ is not available. (Allott, 2007, pg 81) See Figure 4.9, pg 162 In cyclic photophosphorylation, an electron in P700 is excited by a photon and begins taking the same path that it took in noncyclic photophosphorylation. However, the electron is not used to reduce NADP+ but instead is passed back to the b6complex, where the energy is used to generate the proton gradient. The proton gradient is generated in the same manner as in noncyclic photophoshorylation and ATP synthesis by chemiosmosis still occurs. Neither NADPH nor oxygen is produced in cyclic photophosphorylation. (Carter-Edwards, Gerards, Gibbons, McCallum, Noble, Parrington, Ramlochan, Ramlochan, 2011) HOMEWORK: pg 165 #1 - 15 4.2 - Light Independent Reaction (Calvin Cycle) (Taken from Biology 12, MHR, 2011) - occurs in the stroma of chloroplasts This process uses up the ATP and NADPH from light reactions to convert gaseous CO2 into sugar. The Calvin cycle can be divided into 3 phases: carbon fixation, reduction reactions, and ribulose 1, 5-bisphosphate (RuBP) regeneration. See Figure 4.11, pg 166 Phase 1: Carbon Fixation Each CO2 molecule is attached to a 5-carbon sugar called ribulose 1, 5-bisphosphate (RuBP) with the help of an enzyme called ribulose bisphosphate carboxylase/oxygenase (rubisco). This process is called carbon fixation. The result is a very unstable, 6-carbon intermediate that immediately splits into two 3carbon molecules called glycerate-3-phosphate or 3-phosphoglycerate (PGA). The Calvin cycle is also known as C3 photosynthesis (since the first compound to produced contains three carbon atoms). The reaction that leads to these three-carbon compounds can be summarized as: CO2 + RuBP unstable C6 2 PGA Phase 2: Reduction Reactions Each of the 12 glycerate-3-phosphate (or PGA) molecules receive an additional phosphate from an ATP of the light reactions, forming 12 molecules of 1, 3bisphosphoglycerate (1, 3-BPG). A pair of electrons from each of 12 NADPH molecules reduces 12 molecules of 1, 3bisphosphoglycerate to 12 molecules of glyceraldehydes 3-phosphate (G3P). This is a 3C sugar known as triose phosphate (TP). 2 molecules of triose phosphate or G3P exits the cycle as a final product and the remaining ten 3-carbon triose phosphates or G3P’s recycle back towards the regeneration of more RuBP. (These 2 triose phosphate or G3P molecules will be used to make glucose and other sugars.) Phase 3: RuBP Regeneration In a series of enzyme-catalyzed reactions, the remaining 10 molecules of G3P (triose phosphate) are rearranged to regenerate 6 molecules of RuBP to complete the cycle. The Calvin cycle must be completed 6 times in order to make one molecule of glucose. Of the 12 triose phosphate (G3P) molecules produced in six cycles, 10 are used to regenerate RuBP, and 2 are used to make one glucose molecule. It takes 18 ATP and 12 NADPH to produce 6 RuBP and 1 molecule of glucose. Triose phosphate (G3P) is the primary end product of the Calvin cycle. It may be converted into glucose and polymerized into starch within the stroma of chloroplasts, or it may be transported into the cytoplasm and used to produce glucose and sucrose. Sucrose is the main carbohydrate transported from mesophyll cells of the leaf to other cells of the plant. (Carter-Edwards, Gerards, Gibbons, McCallum, Noble, Parrington, Ramlochan, Ramlochan, 2011), (Damon, McGonegal, Tosto, Ward, 2007, pg 231), (Allott, 2007, pg 79) Summary of Photosynthesis The products of the light-dependent reaction (ATP and NADPH) are needed for the lightindependent reaction to occur. Light-independent reaction does not directly need light however it needs the products of the light-dependent reaction. Light-Dependent Reaction occurs in the thylakoids uses light energy to form ATP and NADPH Light-Independent Reaction occurs in the stroma uses ATP and NADPH to form triose phosphate splits water in photolysis to provide returns ADP, inorganic phosphate and NADP + replacement electrons and H , and to release to the light-dependent reaction oxygen to the atmosphere includes two electron transport chains and involves the Calvin Cycle photosystems I and II (Damon, McGonegal, Tosto, Ward, 2007, pg 232) Relationship Between the Structure of the Chloroplast and Its Function Chloroplast Structure extensive membrane surface area of the thylakoids small space (lumen) within the thylakoids Function Allowed allows greater absorption of light by photosystems allows faster accumulation of protons to create a concentration gradient stroma region similar to the cytosol of the cell allows an area for the enzymes necessary for the Calvin cycle to work double membrane on the outside isolates the working parts and enzymes of the chloroplasts from the surrounding cytosol (Damon, McGonegal, Tosto, Ward, 2007, pg 233) Measuring the Rate of Photosynthesis The rate of photosynthesis can be measured directly by measuring the rate of oxygen production or the rate of carbon dioxide intake as long as a correction is made for cell respiration. Production of Oxygen Aquatic plants (e.g., Myriophyllum) release bubbles of O2 when they carry out photosynthesis. If these bubbles are collected, their volume can be measured. Uptake of Carbon Dioxide Leaves take in CO2 from the air or water around them, but this is difficult to measure directly. If CO2 is absorbed from water, the pH of the water rises. This can be monitored with pH indicators or with pH meters. The rate of photosynthesis can also be measured indirectly by measuring the increase in biomass. Increases in Biomass If batches of plants are harvested at a series of times and the biomass of the batches is determined, the rate of increase in biomass gives an indirect measure of the rate of photosynthesis in the plants. (Allott, 2007, pg 21), (Damon, McGonegal, Tosto, Ward, 2007, pg 76-77) Effect of Temperature on Rate of Photosynthesis The effect of temperature on the rate of photosynthesis: Photosynthesis is a biological reaction and like all other such reactions there are steps that require the presence of enzymes. Temperature as we have already met is a change in the average kinetic energy of the particle. The graph the left should look familiar as this is the same one covered in the section on the effect of temperature on the rate of an enzyme catalysed reaction. (a) Increasing rate of photosynthesis as the kinetic energy of reactants increases. 'optimal' temperature. (b) Maximum rate of reaction of photosynthesis at the (c) Decrease in rate of photosynthesis as the enzymes become unstable and denature. (Burrell, J. G. (2002-11) Click4Biology (version 0820.2011). Thailand: Bangkok; URL http://click4biology.info) Effect of Light Intensity on Rate of Photosynthesis The effect of light intensity on the rate of reaction. Light energy absorbed by chlorophyll is converted to ATP and H+ see section 3.8.5. At very low light levels (a) the plant will be respiring only not photosynthesizing. As the light intensity increases then the rate of photosynthesis increases. At high light intensities the rate becomes constant, even with further increases in light intensity there are no increases in the rate. The plant is unable to harvest the light at these high intensities and indeed the chlorophyll system can be damaged by very intense light levels. Effect of CO2 Concentration on Photosynthesis The effect of carbon dioxide concentration on the rate of photosynthesis: Carbon dioxide is one of the reactants of the reaction so this graph is very much like the effect of substrate on the rate of reaction. (a) O2 is used up as the plant is not photosynthesizing but only respiring. (b) As the concentration of the carbon dioxide (substrate) increases the rate of reaction increases. (c) The atmospheric levels of carbon dioxide and the associate rate of photosynthesis. (d) Maximum rate of photosynthesis (see section e). (e) This is a range of values for different plants reaching their saturation level with carbon dioxide. Once the saturation level has been reached there is no further increase in the rate of photosynthesis. (Burrell, J. G. (2002-11) Click4Biology (version 0820.2011). Thailand: Bangkok; URL http://click4biology.info) Alternative Mechanisms of Carbon Fixation C3 Plants (those that use the Calvin cycle to fix carbon) C3 plants use rubisco to add CO2 to RuBP. The resulting 3-carbon molecule is called PGA. Rice, wheat, and soybeans are examples of C3 plants. See Figure 4.12, pg 168 On hot days, most plants close their stomata to conserve water. This decreases the amount of CO2 and increases O2. O2 competes with CO2 for rubisco’s active site. This results in photorespiration, a process where rubisco accepts O2 in place of CO2. When O2 is more plentiful than CO2, O2 binds more often, resulting in the oxidation of RuBP, instead of its carboxylation to PGA. This process is called photorespiration because it occurs in light. Photorespiration decreases the production of carbohydrates by photosynthesis (it removes PGA molecules from the Calvin cycle). Why do plants undergo photorespiration? One hypothesis maintains that this characteristic of rubisco is an evolutionary remnant of an earlier mechanism, which was better suited for an atmosphere rich in CO2 and poor in O2 . Over time, plants did not evolve a modified rubisco or a new fixing enzyme that would bind CO2 and not O2. Some plants, however, have evolved alternative mechanisms of carbon fixation that concentrate CO2 at the site where rubisco is found, thereby suppressing the rate of photorespiration. Two important strategies are C4 photosynthesis and crassulacean acid metabolism (CAM) C4 Plants (undergo C4 photosynthesis) C4 photosynthesis: a photosynthetic pathway of carbon fixation that reduces the amount of photorespiration that takes place by continually pumping CO2 molecules (via malate) from mesophyll cells into bundle-sheath cells, where rubisco brings them into the C3 Calvin cycle. See Figure 4.13, pg 169 C4 plants undergo a different carbon fixation process than C3 plants. Sugar cane and corn are examples of C4 plants. C4 plants have two types of photosynthetic cells; bundle-sheath cells surrounding a vein and mesophyll cells that are located around the bundle-sheath cells. The mesophyll cells are connected to the bundle-sheath cells by cell-cell connections called plasmodesmata (kind of like underground tunnels connecting two buildings). An enzyme called phosphoenolpyruvate carboxylase (PEP carboxylase) catalyzes the addition of CO2 to a 3-carbon phosphoenolpyruvate (PEP), forming the four-carbon molecule oxaloacetate (OAA). This process is called C4 photosynthesis or C4 pathway. OAA is converted to malate. This 4-carbon molecule diffuses from the mesophyll cells into bundle-sheath cells through plasmodesmata. CO2 is removed from the malate molecule (decarboxylation) forming pyruvate (3-carbon) The pyruvate diffuses back into the mesophyll cell where it is converted into PEP The CO2 enters the C3 Calvin cycle in a second fixation reaction, this time catalyzed by rubisco in the bundle-sheath cell. A disadvantage with this process is that an additional ATP is required to regenerate PEP. This method of carbon fixation reduces the amount of photorespiration that takes place by continually pumping CO2 molecules from the mesophyll cells to the bundle-sheath cells (via malate), where rubisco brings them into the Calvin cycle. The high level of CO2 in the bundle-sheath allows the CO2 to outcompete O2 in binding to rubisco. C4 photosynthesis is advantageous in hot tropical climates where photorespiration would otherwise convert more than half of the glucose produced back to CO2. CAM Plants (fix carbon by CAM or crassulacean acid metabolism) Crassulacean acid metabolism (CAM): a photosynthetic mechanism in which stomata open at night so that plants can take in CO2 and incorporate it into organic acids, and close during the day to allow the organic acids to release CO2 molecules that enter the C3 Calvin cycle to be fixed into carbohydrates See 4.14, pg 169 CAM (crassulacean acid metabolism) plants, such as cacti and pineapple plants, open their stomata at night (to gain CO2) and close them during the day to prevent water loss, since they live in very hot, dry, and desert environments. Closing the stomata during the day helps conserve water but prevents CO2 from entering the leaves. In the dark, when stomata open, the plants take in CO2 and incorporate it into C4 organic acids using the enzyme PEP carboxylase. CAM plants incorporate CO2 into an acid called crassulacean acid, within the mesophyll cells and store it in vacuoles overnight. During the day, when ATP and NADPH are made by light reactions, the CO2 is released from the organic acids so it can be incorporated into sugar in the Calvin cycle. In C4 plants, the first part of carbon fixation and the Calvin cycle occur in separate compartments of the leaf (mesophyll cell and bundle-sheath cell). In CAM plants, the two steps occur in the same compartments, but at different times of the day (carbon fixation into organic acids occurs at night and Calvin cycle occurs in the day). The C4 pathway uses almost twice as much ATP as the C3 pathway to produce glucose, however, without this mechanism, photorespiration would reduce the glucose yield to less than half that produced by the C3 pathway. HOMEWORK: pg 171 #1 – 14