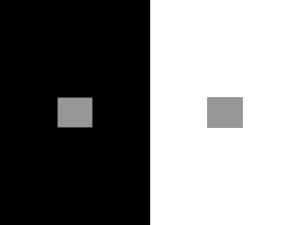P312Ch03B_TheRetina
advertisement

The Retina – the Neural System of the Eye – Chapter 3 G8 Chapter 3 Outline Focusing Light Onto the Retina Light: The Stimulus for Vision The Eye Light is Focused by the Eye Transforming Light Into Electricity How Does Transduction Occur Hecht’s Psychophysical Experiment The Physiology of Transduction Pigments and Perception Distribution of Rods and Cones Dark Adaptation of the Rods and Cones Measuring Cone Adaptation Measuring Rod Adaptation Visual Pigment Regeneration Spectral Sensitivity of the Rods and Cones Measuring Spectral Sensitivity Rod and Cone Absorption Spectra Neural Convergence and Perception Why Rods Result in Greater Sensitivity Than Cones Why We Use Our Cones to See Details Lateral Inhibition and Perception What the Horseshoe Crab Teaches Us About Inhibition Lateral Inhibition and Lightness Perceptioin The Hermann Grid: Seeing Spots at Intersections Mach Bands: Seeing Borders More Sharply Lateral Inhibition and Simultaneous Contrast A Display That Can’t Be Explained by Lateral Inhibition Something to Consider: Perception is Indirect The coverage that follows does not exactly follow the text. The Retina - 1 3/8/2016 Retina: A layer of cells including receptors and neurons on the back of the interior wall of the eye. Neural cells making up the retina include: 1) Rod receptors, 2) cone receptors, 3) bipolar cells, 4) ganglion cells, 5) amacrine cells, 6) horizontal cells. Receptor cells Horizontal cells Bipolar cells Amacrine cells Ganglion cells Light The retina is constructed so that the receptors are farthest away from the light. The light entering the eye strikes and passes through all the other cells and also the bodies of the receptors. Light initiates activity when it strikes the receptor outer segments, the part of the retina furthest from the light. Why do the receptors face away? Nutrition. It’s more efficient to supply the nutrient-carrying blood to the receptors from the back of the eye. The Retina - 2 3/8/2016 The receptors Two types – Rods and Cones What the receptors have in common 1. They all transduce light into release of neurotransmitter substance. In the absence of light, both rods and cones release glutamate, a neurotransmitter substance. When light strikes them, they reduce the amount of glutamate released. So, light is signaled by a reduction in release of neurotransmitter substance. 2. All receptors become more sensitive to light when placed in darkness and less sensitive when placed in the light. The change in sensitivity occurs over several minutes. 3. They all synapse with bipolar cells. Differences between rods and cones 1. Location: Cones are located in the center of the retina; rods are in the periphery. 2. Sensitivity: Cones are much less sensitive to light than are rods. So, at night, we are better able to see faint lights out of the corner of our eyes. Looking directly at a faint light in the night may make it disappear. Rods quit working when light level is about that corresponding to twilight. They’re “max’d” out. Cones require low sunlight or better to be activated. This graph does not do justice to the huge 1000:1 difference in sensitivity between the cones and rods. Sensitivity Cones Rods The Retina - 3 3/8/2016 3. Number: There are about 6 million cones, about 100 million rods per eye. 4. Types: There are 3 types of cones, only one type of rod. 5. Differential sensitivity to wavelength. G8 p56 Let’s name the cones, the S, for short wavelength, M, for medium wavelength, and L for long wavelength cones. The S receptors are actually sensitive to ultraviolet radiation, but lens absorbs UV radiation, so none of it strikes the receptors. 6. Differential wiring: Differential connections to bipolar and ganglion cells. Cones are more likely to have few-to-1 connections – few cones, perhaps only one, drives one bipolar which drives one gangliion cell. Rods are characterized by many-to-1 connections – 100s, sometimes 1000s of rodsx synapse with one bipolar. Many of these bipolar cells synapse with 1 ganglion cell. So: Cones: few-to-1 Rods: many-to-1 The Retina - 4 3/8/2016 Questions to consider 1. Why two types of receptor? Why not just one type? One outfielder cannot cover the whole outfield, so baseball has left fielders, center fielders, and right fielders. My guess is that the range of intensities to which we’re sensitive is too large to be covered by just one receptor type. Two receptor types – one for night and one for day provides better coverage of the range of lights in which we have to operate. Range of intensities in which we exist is about 1 million to 1. That is, the brightest sunlight in which we are able to use our eyes is about 1 million times more intense than the faintest starlight in which we can still see shadows. But the range of responses of receptors and the bipolar – ganglion cells to which they connect is only about 500-1000 to 1. This means that significant changes in intensity would be represented by very small changes in response rate of the cells involved. This would likely result in many intensity changes going unnoticed. So the two receptors “divide” up the intensity range, with the rods taking the faint lights and the cones taking the bright lights. 2. Why 3 types of cone? Why not just one type? The short answer is that having three types of cones makes it MUCH easier for us to identify different wavelengths than if we had just one. Review the previous lecture on the simple circuit to detect short and long wavelengths. If we had only one receptor type, the world would be only shades of gray. The fact is that the more cone types you have, the better able you are to discriminate wavelengths – the richer your color perception is. See figures on next page. More on this in the color perception chapter. 3. Why don’t we have more than 3 types of cones? Wouldn’t our color vision be even better than it is now? The answer is yes. However it turns out that each type of cone requires many thousands of extra neurons to process and integrate the information from that type. But the brain has other things to do than to provide good color vision. What about music? Form? Touch? Arithmetic? Literature? So the choice is to devote extra neurons to better color vision or devote them to processing auditory information, or form, or touch, or something else. The bottom line is that our brain is a compromise. More on this in the chapter on color The Retina - 5 3/8/2016 Perceptual consequences of our having two types of receptors 1. Dark Adaptation G8 p52 When put in the dark, our sensitivity to light increases, then levels off after about 2 minutes, then increases again for up to 30 minutes. Measuring dark adaptation. 1) Expose the participant to light for > 5-10 minutes. 2) Put the participant in an absolutely lightless environment. 3) Wait a predetermined time, then quickly determine the threshold for detection of light. Repeat over and over using different times in the dark. Plot the threshold as a function of time in the dark. Threshold – lower means more sensitive 3 min 20-30 min Time in dark before threshold measured Difference in thresholds is about 1,000,000 to 1 between Time 0 and 30 minutes in the dark. Our visual system is about 1 million times more sensitive after 30 minutes in the dark. Early researchers thought the “bump” in the curve was an artifact. But they kept getting the bump. Probably some research assistants were fired because their superiors thought they were careless. We now know that the upper part of the curve is the dark adaptation curve of cones and that the lower part of the curve is the dark adaptation curve of rods. Why did pirates wear eye patches? 2. The Purkinje shift – another consequence of two receptor types. G8 p. 57 As it gets dark, blue-green objects become brighter relative to other objects than they are when it’s light. When it’s dark outside, the rods contribute much to our vision. Because they’re most sensitive to light that’s blue-green, those colors appear brighter relative to others when it’s dark. This shift in brightness is called the Purkinje shift. When it’s bright outside, the rods contribute little to vision, and because the cones, as a set, are not as responsive to blue-green, blue-green objects don’t seem overly bright in high illumination. Rods can’t see red, so emergency equipment that needs to be seen at night should not be red. The best compromise is a bluish/green combination – visible by both rods and cones. 3. Different parts of the retina are used for different purposes. Center for fine detail in lighted conditions – the wiring of the cones Center for color perception – the 3 types of cones Periphery for faint lights – the rods are more sensitive and wired for sensitivity. The Retina - 6 3/8/2016 Other cells in the retina. We’ll first follow the neural activity upwards toward the brain – along the receptor -> bipolar -> ganglion pathway. The Bipolar cells. G8 p59 So called because it’s difficult to tell which is the “receiving” end and which is the “sending” end. They transmit information from the receptors to the ganglion cells. Two types: One responds positively to increase in glutamate. Recall that glutamate is released in abundance in the absence of light. Thus, this converts absence of light into a positive response. Others respond positively to decreases in glutamate. This converts presence of light into a positive response Why do they exist? Perhaps to provide an extra synapse between the receptors and ganglion cells.?? The first synapse, the receptor –> bipolar synapse may allow one type of comparison, perhaps local. That is, other cells beside the receptor also synapse with the bipolar cell, resulting in comparisons. This is called lateral inhibition. The second synapse, the bipolar –> ganglion synapse may allow a completely different type of comparison, perhaps more global. This is pretty much the way things are in the brain – the comparisons, decisions, “thinking”, of the brain is done in the synapses. The Retina - 7 3/8/2016 Ganglion cells Only cells in retina that are clearly neurons. About 1 million per eye. Axons form the optic nerve. So EVERYTHING WE SEE from an eye is carried to the rest of the brain over the 1 million axons of the ganglion cells in that eye. This is a 100:1 compression of information. Ganglion cells receive neurotransmitters from bipolar and amacrine cells. Most of them send their axons to the lateral geniculate nucleus. Their axons are a few inches long. Three general types of ganglion cell: M for Magni, larger in size with large receptive fields. P for Parvi, smaller with smaller receptive fields. K for koniocellular, kind of like P cells, with B/Y antagonism. The M ganglion cells apparently carry information about movement and about gross features of the visual environment. The P ganglion cells apparently carry information about wavelength and about fine details of the visual environment. K cells - not well understood right now. The blind spot The axons of the ganglion cells cross the surface of the retina and gather together at one place. There, they go toward the back of the head, out of the eye. That spot, obviously, has no receptors. It is called the blind spot. G8 p 51-52 with a demo of the blind spot on p. 52. Five of the approximately 1 million ganglion cells Blind spot The Retina - 8 3/8/2016 The Gossiping cells of the retina Horizontal Cells These cells receive input from receptors and send inhibition to neighboring bipolar cells. Since the inhibition is sent laterally, it’s called lateral inhibition. R B R B H B R R Lateral Inhibition One likely function of the horizontal cells is to heighten the contrast of borders between light and dark. The amacrine cells These cells receive input from bipolar cells and send inhibition to neighboring ganglion cells. G G A Amacrine B High level Lateral Inhibition B Horizontal H R R One conjecture is that the Amacrine cells are involved in inhibition associated with wavelength, such as illustrated in the previous lecture on use of wavelength information. Amacrine cells may act as switches between rod and cone systems driving the same ganglion cell. The Retina - 9 3/8/2016 Lateral Inhibition – inhibition that is propagated across the retina. G8 p61 The transmission of inhibition laterally from one part of the retina to another adjacent area is called lateral inhibition. Lateral inhibition plays a very important part in the processing of the visual image in the retina. It is apparently involved in the following aspects of that processing . . . 1) Antagonism between center and surround of ganglion cell receptive fields. 2) Heightening or sharpening contrasts between adjacent light and dark areas. 3) Illusions such as Mach bands and the Hermann grid. 4) Simultaneous contrast 5) The “creation” of colors from the responses of the three cone types. The Retina - 10 3/8/2016 Phenomena explained (at least partially) by lateral inhibition 1) Mach Bands Actual Intensity Perceived Intensity The Retina - 11 3/8/2016 Text Figure 3.37, p. 65 Light Receptors 100 -10 100 -10 -10 100 -10 -10 20 -2 -10 20 -2 -2 Horizontal cells 20 -2 -2 -2 -2 Bipolars 100-10-10 = 80 100-10-10 = 80 100-10-2 = 88 20-10-2 =8 The Retina - 12 20-2-2 = 16 20-2-2 = 16 3/8/2016 2) Hermann Grid The Hermann Grid is illustrated in G8 p 62-64. Below is an example of the figure. Most persons see gray dots in the intersection of the lines in the periphery of their vision, although they do NOT see the dots in the center of their visual field. The explanation is based on the fact that receptive fields in the periphery are larger than those in the fovea. Perception in the periphery: Receptive fields are larger than the streets and intersections. Most of surround Peripheral perception of the “intersection” is darker than in the light so the perception of the “street” because surrounds of the ganglion cell receptive fields of “intersection” ganglion cells are response is “gray” mostly in the light, inhibiting the responses of those Fixation ganglion cells. Peripheral perception of the “street” is lighter than of the “intersection” because surrounds of receptive fields of those ganglion cells are mostly in the dark, resulting in less inhibition. point Most of surround in the dark so ganglion cell response is “light” Perception in the fovea: Receptive fields are small, so the whole receptive field lies within the street and perception remains the same regardless of location. In the center of the visual field, the receptive fields are so small that the whole receptive field is in the light regardless of whether you look at the intersection or the middle of the “street”. The Retina - 13 3/8/2016 3) Simultaneous Contrast (To demonstrate, grab one of the squares and hit the Backspace key. Then press CTRL+Z to undo.) The circles are all EQUALLY intense. The explanation is that when the surround is bright, that brightness inhibits the responses of ganglion cells perceiving the center, making the center seem darker. The Retina - 14 3/8/2016 Sensitivity vs. Acuity G8 p58 Convergence ratio: The number of receptors “connecting” to a single ganglion cell. Low convergence ratio: Just a few receptors “connect” to a ganglion cell. High convergence ratio: Many receptors “connect” to a ganglion cell Resolution: Ability to distinguish bands of dark or light. High resolution/acuity: Ability to distinguish narrow bands Low resolution/acuity: Only able to distinguish wide bands The relationship of resolution to convergence ratio High convergence ratios -> Low resolution/acuity Response is the same regardless of where the dark bar is located. Low convergence ratios -> High resolution/acuity A A responds B A The Retina - 15 B B responds 3/8/2016 Convergence ratio and sensitivity Sensitivity: Ability to detect faint lights. High sensitivity: Good ability to detect faint lights Low sensitivity: Ability to detect only bright lights High convergence ratios -> High sensitivity Responds to faint light Low convergence ratios -> Low sensitivity A B Neither responds The Retina - 16 3/8/2016 Obtaining Good acuity and high sensitivity in the same overall system. How does the visual system handle the dual demands of acuity and sensitivity? To have good acuity, we need small receptive fields, with 1:1 connections between receptors and ganglion cells. To have high sensitivity, we need large receptive fields with many:1 connections between receptors and ganglion cells. Apparently, a two-pronged solution was required.. . . 1. Receptors with different individual sensitivities. 2. Subsystems with different convergence ratios. These are combined to form two separate systems. The scotopic system: Rods with high individual sensitivity and high convergence ratios. The photopic system: Cones with lower individual sensitivity and low convergence ratios. The retina could have been all cones connected to ganglion cells with different convergence ratios. But cones are not individually sensitive enough. The retina could have been rods and cones, with equal convergence ratios But increasing convergence is needed to maximize sensitivity. The duplex solution in the eye is a wonderful compromise. The Retina - 17 3/8/2016 Possible test question concerning dark adaptation. You take your nephew or niece to a movie. When you enter the theater, you can barely even see the seats. Eventually you are able to see much better than just after you’d entered. Your nephew or niece asks you, “Why can I see so much better now than when we just came into the theater?” What are the reasons that after 30 minutes in darkness you are so much more sensitive to light than when you just came into the theater. 1. Individual cones have become more sensitive if they’re in the dark for a period of time – due to changes in the molecules within the cones. 2. The visual system begins using output from rods, each of which is individually more sensitive to light than cones. 3. The visual system beings using output from rods, which are wired for spatial summation, increasing sensitivity. 4. Individual rods have become more sensitive after being in the dark for a period of time – their internal chemicals – retinal and opsin – have recombined. The Retina - 18 3/8/2016 Review of issues associated with the neural apparatus of the eye Issues associated with receptor placement – facing forward vs. facing backward Issue of number of receptor types – one vs. two or more Issue of differences in receptor types – difference in sensitivity, difference in preference for wavelength Issues of number of cells in the eye – receptors vs. receptor + ganglion vs. receptor + bipolar + ganglion. Issues of how ganglion cell receptive fields should be organized. Issues of convergence ratio and acuity vs. sensitivity. The Retina - 19 3/8/2016