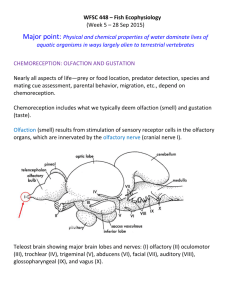
fulltext
advertisement

Lateralization of Human Olfaction: Cognitive Functions and Electrophysiology Daniel A. Broman Department of Psychology Umeå University, Sweden 2006 All my genius is in my nostrils - Nietzsche Copyright © 2006 Daniel A. Broman ISBN 91-7264-166-5 Printed by Solfjädern Offset AB, Umeå II ABSTRACT Broman, D.A. (2006). Lateralization of human olfaction: cognitive functions and electrophysiology. Doctorial dissertation from the Department of Psychology, Umeå University, SE-90187, Umeå, Sweden: ISBN 91-7264-166-5 In this thesis lateralization of olfactory functions was investigated by both behavioral and electrophysiological assessment, the latter with the olfactory event-related potential (OERP) technique. The olfactory sense is primarily ipsilateral in that a stimulus that is presented to one nostril is initially processed in the same hemisphere. This makes it possible to observe differences between stimulated nostrils as an indication of hemispheric difference. Study I explored differences in olfactory cognitive functions with respect to side of rhinal stimulation and demonstrated that familiarity ratings are higher at right- compared to left-nostril stimulation. No differences were found in episodic recognition memory or free identification, possibly reflecting inter-hemispheric interactions in higher cognitive functions. Effects of repetition priming were present in odor identification and tended to be more pronounced when tested via left nostril. Study II further investigated the effect of previous exposure in odor identification by a different experimental set-up, and demonstrated effects of repetition priming when tested via left- but not right-nostril stimulation. This finding indicates the importance of reconsidering possible sequential effects in olfactory research. Study III examined methodological aspects of an OERP protocol with respect to stimulus duration, which was used in Study IV. No differences in amplitudes or latencies where found between the stimulus durations of 150, 200 and 250 ms, suggesting the commonly used duration of 200 ms in a standard protocol. Study IV investigated laterality effects in OERPs with respect to side of stimulation and electrode site. The results showed consistent amplitudes and latencies regardless of rhinal side of stimulation. Larger amplitudes were demonstrated on left hemisphere and midline compared to right hemisphere, possibly explained by smaller N1/P2 amplitudes at the right-hemisphere sites at left-nostril stimulation. Apart from a proposed OERP protocol, the findings support the notions of a righthemisphere predominance in processes related to olfactory perception and indicate, in accordance with other findings, a left-side advantage in conceptual repetition priming. KEY WORDS: Human olfaction, Laterality, Olfactory processes, Odor familiarity, Odor identification, OERP III ACKNOWLEDGEMENTS First and foremost I sincerely want to thank my supervisor Dr. Steven Nordin. I am also deeply indebted to Dr. Mats J. Olsson. Further, I want to thank my other co-authors, with a special acknowledge to Jonas Olofsson as first author on the fourth article included in the thesis, Dr. Lars Nyberg and Lena Öman for valuable comments on the thesis, and Dr. Gregory Neely for valuable comments on manuscript. Moreover, thanks to Dr. Claire Murphy and her group at the Life-Span Human Senses Laboratory, SDSU/UCSD, San Diego, USA and Dr. Per Møller and his group at the Sensory Science Laboratory, KVL, Copenhagen, Denmark, for inviting me to visit their laboratories and conducting olfactory research as a pre-doctorial fellow. Finally, I would like to thank the Kempe Foundation, the Sweden-America Foundation and the Wallenberg Foundation for financially supporting me and my research. IV LIST OF PAPERS This doctorial thesis is based on the following studies: I. Broman, D.A., Olsson, M.J. & Nordin, S. (2001). Lateralization of olfactory cognitive functions: effects of rhinal side of stimulation. Chemical Senses, 26, 1187-1192. II. Broman, D.A. & Wising, A. (submitted). Effects of repetition priming and stimulated nostril on odor identification. III. Broman, D.A., Nordin, S., Gilbert, P.E. & Murphy, C. (submitted). Olfactory event-related potentials from varying stimulus duration in young and elderly adults. IV. Olofsson, J.K., Broman, D.A., Gilbert, P.E., Dean, P., Nordin, S. & Murphy, C. (2006). Laterality of the olfactory eventrelated potential response. Chemical Senses, 31, 699-704. V VI TABLE OF CONTENTS INTRODUCTION AND BACKGROUND Anatomy of the human olfactory system Research methods in human olfaction Behavioral testing Methods measuring neural activity Olfactory lateralization Absolute sensitivity Discrimination Recognition memory Identification Neural activity 1 2 3 3 5 8 9 9 10 10 11 RESEARCH OBJECTIVES 12 SUMMARY OF EMPIRICAL STUDIES Study I: Lateralization of Olfactory Cognitive Functions: Effects of Rhinal Side of Stimulation Study II: Effects of Repetition Priming and Stimulated Nostril on Odor Identification Study III: Olfactory Event-Related Potentials from Varying Stimulus Duration in Young and Elderly Adults Study IV: Laterality of the Olfactory Event-Related Potential Response 14 14 16 18 19 GENERAL DISCUSSION Behavioral findings ERP findings Lateralization of olfactory function Conclusions 21 21 22 22 27 REFERENCES 29 VII VIII INTRODUCTION AND BACKGROUND Historically the chemical senses have, compared to the other senses, not been extensively investigated. Although scientific studies have been investigating aspects of perception and cognition in the olfactory sense for more than a century (e.g., Fischer & Penzoldt, 1886; Toulouse & Vaschide, 1899; Zwaardemaker, 1895), the extent of work in this field is modest in relation to research in vision and audition. In some aspects this may perhaps seem natural since vision and audition are more related to several higher cognitive functions, such as speech communication and reading. The understanding for these sensory systems and their basic transduction is also deeper. It is only a couple of years ago that the chemical senses received their first Noble Prize for findings concerning the olfactory receptors (Buck & Axel, 1991, for the subsequent work see, for example, Buck 2000; Buck 2004). However, somewhat intriguing, the relation between molecular structure and the perceived odor is still not clearly understood. For example, the imaginative theory that the perception of a certain odorant is determined by molecular vibrations coded by the receptors (Turin, 1996) had some bearing until recently when it was experimentally tested with no support for it (Keller & Vosshall, 2004). In addition to the aspects mentioned above, one reason for odor perception and cognition not having been as vividly explored as vision and audition is probably related to difficulties associated with the stimulus. Visual and auditory stimuli are in many aspects easier both to produce and present with superior control. They are also less complicated to measure in physical units, to vary in a single dimension (e.g., frequency or amplitude) and to categorize. Odor categorization is a good example of possible caveats related to stimuli. Several attempts have over the years been made to find good categorization systems (e.g., Linnaeus, 1756; Henning, 1916; Amoore, 1963). However, as noted by Doty (1991), odor categorization is still enigmatic and classification schemes focused on explaining olfactory function have, more or less, fallen out of favor in the past decades. One reason for the lack of a classification system related to odor perception that is generally agreed on is probably that the odor information to a large extent is based on experience and therefore also individually tuned. For issues related to this, see Hudson (1999) and Wilson and Stevenson (2003). Recently, results from an imaging study indicate different coding of odor quality and odorant structure in human piriform cortex, which may explain why odor quality 1 to some extent can be independent of simple odorant structure (Gottfried, Winston & Dolan, 2006). At first inspection, it is easy to believe that since the olfactory sense not is involved in higher-cognitive function in the same way as the visual or auditory sense it is also less optimalized in humans. Trygg Engen (1991) would probably argue for the opposite, based on its purpose this sense is extremely well suited. We may not learn new languages or solve mathematical problems with assistance from the olfactory sense, but we certainly make up our minds about what to eat, and not to eat, on bases of the olfactory system and can also detect the smell of a fire. Anatomy of the human olfactory system In humans there are two chemical senses with receptors present in the nose that have been proved to contribute to our perception of odorants (e.g., Cain, 1974; Cometto-Muñiz & Cain, 1990; Doty, Brugger, Jurs, Orndorff, Snyder & Lowry, 1978). In addition to the transduction of odor information by the olfactory nerve (CN I), chemo-somatosensory information including irritation and pungency is mediated by the trigeminal nerve (CN V). Most odorants that we are exposed to are also, at least at higher concentrations, to some extent trigeminal (Doty, 1975). Interestingly, when an odor without trigeminal properties is monorhinally presented it is not possible to tell whether it is presented to the left nostril or to the right. To be able to localize nostril of stimulation the odor has to have trigeminal properties (Kobal, van Toller & Hummel, 1989; Radil & Wysocki, 1998). However, for a different view see a recent neuroimaging notion about spatial information abilities (Porter, Anand, Johnson, Khan & Sobel, 2005). The olfactory nerve (CN I) is formed by the axons of millions of receptor cells in the olfactory epithelium. The cilia of the receptor cells are distributed in the nasal mucosa in the upper part of the cavity of each nostril and respond to chemical stimulation. Different types of receptors code for different odorants depending on their chemical structure. The axons of the receptor cells traverse the cribriform plate and extend into glomeruli in the olfactory bulb. The olfactory bulbs are located on the lower surface of the frontal lobes, and each receptor projects ipsilateral to one glomerulus in the bulb. The glomeruli contain dendrites of mitral and tufted cells. These second-order neurons project via the lateral olfactory tract directly to the primary olfactory cortex of each hemisphere (Price, 1990). 2 The primary olfactory cortex consists of the anterior olfactory nucleus located in a dorsal segment of the olfactory bulb, piriform cortex and entorhinal cortex. Close to these areas, and with several connections, we also find the hippocampus and the amygdala (McLean & Shipley, 1992). Olfactory information is later transferred to parts of neocortex, including oritofrontal cortex, by direct projections from primary olfactory cortex as well as by connections via thalamus (Doty, Bromley, Moberg & Hummel, 1997). By first inspection, the earliest inter-hemispheric connection would be by the medial olfactory tract. However, this structure does not seem to be developed in humans (Price, 1990). Thus, possible projections between the hemispheres with respect to odor information are via anterior commisure, with few contralateral connections with respect to the olfactory system (Shipley & Ennis, 1996), and later via corpus callosum. Accordingly, initial olfactory processing in the main olfactory system is primarily ipsilateral. Research methods in human olfaction Behavioral testing There are numerous ways to assess olfactory functions by means of behavioral testing. The tests vary from measuring perceptual functions, such as odor detection, to, for example, odor recognition memory or odor identification that also tap on higher cognitive processes. Absolute detection, the lowest concentration of an odorant possible to detect, is commonly studied by the psychophysical method of constant stimuli or method of limits (Engen, 1971). In the method of constant stimuli a set of stimuli is presented repeatedly in a randomized order. The data obtained can be displayed as a psychometric ogive function, and the threshold is often defined as the stimulus concentration that can be detected in 50% of the cases. The method of limits is less time consuming, but can also be less precise. This method starts with a stimulus that is clearly above or below the absolute threshold and thereafter presents stimuli in descending or ascending series with small intervals until the absolute threshold is reached. The staircase procedure is a variant of this method that shifts the series order when the threshold is reached (Gescheider, 1997). Similarly, these methods can be used for studying odor discrimination with respect to both intensity and quality. However, the most common way to assess discrimination abilities is by 3 making direct comparisons between stimuli. One way is by same/different judgments after inspection of odor pairs, another is by finding the different stimulus out of three. There are different ways to test odor identification. The most complex and difficult type of identification is probably by veridical naming, i.e., correct verbal naming of an odorant without any further information. An easier type of task is to identify an odor with provided alternatives, a so-called multiple-choice test. The typical way to study episodic memory in olfaction is by testing recognition memory. The test consists of an encoding phase were the participants are presented to a set of odorants followed by a retrieval phase, including presentation of the encoded odors to be recognized (targets), and previously not encoded odors (lures). Perceived properties of an odor stimulus, such as intensity, valence and familiarity, can also be rated. One common psychophysical method is by means of magnitude estimation, most frequently used for intensity ratings. The relation between stimulus proportion and the perceived magnitude can be described by the psychophysical power law presented by S.S. Stevens (1957). Another way to measure the magnitude of the percept is by using scales. Both numerical scales with verbal anchors, for example the category-ratio scale (Borg, 1982; Borg, 1998; Borg & Borg, 1998) and the label-magnitude scale (Green, Shaffer & Gilmore, 1993; Green, Dalton, Cowart, Shaffer, Rankin & Higgins, 1996), and the visual-analog scale (Dexter & Chestnut, 1995; Neely, Ljunggren, Sylven & Borg, 1992) are considered to provide measurement on a ratio-scale level. Even if the majority of olfactory research is conducted more or less by non-standardized stimuli and assessments, there is an increase of standardized olfactory tests available for different purposes. For example, the University of Pennsylvania Smell Identification Test (UPSIT; Doty, Shaman & Dann, 1984), the San Diego Odor Identification Test (SDOIT; Andersson, Maxwell & Murphy, 1992), the Scandinavian Odor Identification Test (SOIT; Nordin, Brämerson, Lidén & Bende, 1998), all of these identification test with forced-choice alternatives. Other tests, measuring various functions, are the Connecticut Chemosensory Clinical Research Center Tests (CCCRC; Cain, 1989) for absolute detection and identification, and the Sniffin’ Sticks (Hummel, Sekinger, Wolf, Pauli & Kobal, 1997) measuring absolute detection, intensity, quality discrimination, and identification. 4 Methods measuring neural activity During the last two decades the use of methods measuring neural activity has dramatically grown in human olfactory research, especially the technique of ERP and the imaging techniques of PET and fMRI. For pioneering work using the techniques of ERP and neuroimaging in olfactory research, see Kobal (1981) and Zattore and colleagues (Zatorre, Jones-Gotman, Evans & Meyer, 1992), respectively. These research methods are powerful tools for the study of human olfaction and can naturally also provide useful information concerning lateralization of olfactory functions. Event-related potentials Event-related potentials (ERPs) reflect electrophysiological activity evoked by a stimulus. For a single recording this neural electrical activity is a relatively small section of the registered electroencephalogram (EEG) that is mainly caused by non-stimulus related activity. By computing the mean electrical activity from repeated stimulus presentations with a timelocked stimulus onset, the noise ratio in the event-related activation will be reduced and provide an electrophysiological waveform related to the stimulus and task of interest (Handy, 2005). In this wave, reflecting electrophysiological activity elicited by a specific stimulus and time frame, components can be identified as positive or negative peaks in relation to a pre-stimulus baseline. The peaks are commonly named by their positive/negative deflection and position in relation to stimulus onset. For example, P1 for the first positive peak, N1 for the first negative peak, P2 for the second positive peak, N2 for the second negative, and P3 for the third positive peak. See Figure 1. The peaks of interest to interpret for a specific research question depends on stimulus modality, task given to the participant, study design and research questions (Polich, 2003). Latencies of different types of peaks are closely related to the stimulus modality. For example, in audition N1 and P3 are sometimes referred to as N100 and P300, respectively, because of their approximate presence in ms after stimulus onset. 5 30 P3 Amplitude (µV) 20 P2 10 0 -1 0 N1 -2 0 0 200 400 600 800 1000 1200 1400 1600 L a te n c y (m s ) Figure 1. Example of an ERP registration with the N1, P2 and P3 components inserted (noteworthy, the P3 component has typically a lower amplitude). In olfactory event-related potentials (OERPs) the component of a possible P1 is rarely studied due to the small and unreliable amplitude, neither has there been extensive focus on N2 in OERPs. The main components of interest in OERP research have instead been N1, P2 and P3, and in the most studies amplitudes and latencies of these peaks and their relation to each others have been investigated (e.g. N1/P2, peak-topeak difference). The early components, in olfaction studied by N1, and in some cases also by P2, are often referred to as sensory components that reflect more exogenous aspects in relation to the stimulus (Kobal, 2003). Later peaks, especially P3 and to some extent P2 (which in olfaction sometimes together with P3 constitutes a late positive complex rather than isolated peaks), reflect more endogenous components of more cognitive nature. OERPs can be recorded from electrodes placed on any position on the scalp. Usually, the recording sites follow the International 10-20 system with origin points from nasion, inion and prearciulars (Evans, Kobal, Lorig & Prah, 1993). Based on these points, electrodes are placed along the saggital and coronal plane. Following the midline (z=0) in the saggital plane, the site labelled Cz (central zero) is of equal distance to nasion and inion, the site Fz (frontal zero) is located 20% of the distance between nasion and inion anterior of Cz, and the site Pz (parital zero) is at the same distance posterior of Cz. In the coronal plane, following the central axis, the main points from left to right are labeled 6 T3, C3, Cz, C4, T4 (i.e., odd numbers refer to the left hemisphere and even numbers to the right hemisphere), with similar distance distribution as for the points in the saggital plane. On basis of these main axes, other points are given, for example F3 (frontally on left hemisphere) placed perpendicular to Fz and C3. See Figure 2. Figure 2. Recording sites, according to the International 10-20 system, used in Study III (only midline sites: FZ, CZ, PZ) and Study IV. Haemodynamic techniques With functional neuroimaging it is possible to investigate brain activity with good and reliable spatial resolution. Positron emission tomography (PET) commonly measures local changes in blood flow (Raichle, 1994). When a specific area is more activated there is an increase of regional cerebral blood flow (rCBF), and this increase in rCBF is measured by a tracer, such as 15O. The temporal resolution in this technique is rather poor and the experimental set-up is limited to block design. Neural activity measured by local differences in blood flow can also be studied by functional magnetic resonance imaging (fMRI). Changes in rCBF are with this method studied through differences in blood oxygenation, and the so-called blood oxygen level dependent (BOLD) signal is measured by a magnetic resonance scanner (Ogawa, Lee, Kay & Tank, 1990). In comparison to PET, one advantage of fMRI is the better temporal resolution that also permits event-related designs, even though these designs are, hitherto, not commonly used to study olfaction. Other advantages include fMRI being non-invasive and less expensive. A major disadvantage of fMRI in olfactory research is the presence of artefacts in the signal in areas consisting of air and tissue. Although there are methods to reduce this type of artefact located in olfactory regions in 7 ventromedial prefrontal cortex (Liu, Sobering, Duyn & Moonen, 1993; Yang, Darzinski, Eslinger & Smith, 1997; Zald & Pardo, 2000), the signal quality in this area is only moderate (Royet & Plailly, 2004). Stimuli presentation is often a methodological issue in olfactory research and the use of imaging techniques makes no exception to that. Besides adaptation and habituation problems in the commonly used block design, and administration problem in the scanner, effects of sniffing behaviour and baseline contrasts contribute to somewhat conflicting imaging findings. The most striking inconsistency concern registered activation in the primary olfactory cortex. Although, the signal artefacts in this region in some cases may contribute to the discrepancies in the fMRI results, this is obviously not the whole explanation, since PET studies also show inconsistency in finding activation in the primary olfactory cortex. It has previously been well demonstrated that sniffing behaviour affect olfactory processing (Laing, 1983; Teghtsoonian, Teghtsoonian, Berglund & Berglund, 1978). However in relation to imaging, Sobel and colleagues (Sobel at al., 1998, Sobel et al., 2000) reported that sniffing odorless air elicited activation in primary olfactory cortex. Consequently, the use of that type of task as baseline condition for contrast analysis can explain the lack of activation in some imaging results. Possible effects of adaptation and habituation have also been suggested explanations for the different results in finding primary olfactory cortex activation (Poellinger et al. 2001; Sobel et al., 2000). To conclude, lack of demonstrated activation in primary olfactory cortex in olfactory imaging results should not be interpreted as lack of involvement of primary olfactory cortex in olfactory processing. Olfactory lateralization The fact that the initial processing in the olfactory system is principally ipsilateral to the stimulated nostril, and the direct projections to olfactory related areas in cortex, provide a unique opportunity among the senses to investigate lateralization differences by means of behavioral testing. A great deal of the knowledge about olfactory lateralization originates from studies in patient groups with cerebral complications, either by comparing left- versus right-nostril performance, or by comparing patients with left- versus right-hemisphere lesions. More recently, new and better techniques to study neural activity have been developed that provide further understanding of lateralization of olfactory function, and some of these findings will also be discussed below. 8 Absolute sensitivity Studies that have investigated differences in absolute sensitivity between left- and right-nostril stimulation with behavioral measurements have in many cases shown somewhat conflicting findings. For odor-detection thresholds, Youngentob and associates (Youngentob, Kurtz, Leopold, Mozell & Hornung, 1982) reported differences between the nostrils, where left-handed participants were more sensitive in the left nostril and right-handed participants tended to be more sensitive in the right nostril. In contrast, Cain and Gent (1991) found right-nostril superiority in detection sensitivity, irrespective of handedness. In addition, a study of Koelega (1979) failed to find any differences in thresholds between nostrils. Furthermore, no side differences regardless of handedness were found by either Zatorre and Jones-Gotman (1990) or Betchen and Doty (1998). Interestingly, these two studies had the largest sample sizes of those cited above, and both used phenyl ethyl alcohol (PEA), an odor with limited trigeminal properties (Doty et al., 1978). To conclude, if differences between the nostrils in absolute sensitivity do exist, the size of this effect is probably rather small, and the results suggest a right-nostril superiority, at least for right-handed individuals. Discrimination In discrimination, in accordance with absolute sensitivity, the findings are somewhat conflicting, but they seem to indicate a similar right-nostril superiority. In investigating odor quality discrimination, Zatorre and Jones-Gotman (1990) reported a right-nostril advantage irrespective of handedness. In line with that finding, a right-nostril advantage has also been reported by Martinez and colleagues (Martinez, Cain, de Wijk, Spencer, Novelly & Sass, 1993). However, Hummel, Mohammadian and Kobal (1998) reported that left-handed participants had a left-nostril advantage for this task. In a study by Savic and Berglund (2000), a significant right-nostril superiority in quality discrimination was present only for unfamiliar, but not for familiar odors. Interestingly, inspection of the presented results in Table 2 of that study reveals that, nominally, the right-nostril advantage is present also for familiar odors, which raises the question whether the lack of a significant difference in this case can be related to ceiling effects. A right-nostril advantage has also been shown in an intensity categorization task, which also taps on memory functions, 9 although these findings were statistically significant only for women (Pendense, 1987). Recognition memory Studies of odor recognition memory have reported hemispheric differences in patient groups. Several investigations (Abraham & Mathai, 1983; Jones-Gotman & Zatorre, 1993; Rausch, Serafetinides & Crandall, 1977) have shown that patients with right-temporal lobe lesions perform more poorly than patients with left-temporal lobe lesions in odor recognition tests. This somewhat consistent finding may suggest a righthemisphere superiority in odor recognition. Further, investigating healthy participants in a cross-modal recognition task (odor-word, odor-picture) with the odor as the first stimulus presented birhinally, the response time was faster when the second stimulus (word or picture) was presented to the right hemisphere compared to the left hemisphere (Zucco & Tressoldi, 1989). Using a unimodal recognition task, Olsson and Cain (2003) found no differences in memory performance between nostrils, although the response latency was shorter when tested on the right-nostril compared to the left. However, in a study by Bromley and Doty (1995) the results showed no evidence for a right-nostril advantage in recognition memory, using either a single or a multiple target test. In accordance, Annett, Ford and Gifford (1996) found no side differences in odor recognition without verbal elaboration in an experiment with a birhinal retrieval phase. Identification Gordon and Sperry (1969) found that patients with surgically disconnected hemispheres are able to identify odors verbally when presented to the left but not the right nostril, which could be explained by a left-side lateralization for language. Noteworthy, using non-verbal tests the patients were able to identify odors presented to the right nostril. However, there are indications of a left-hemisphere advantage in odor identification in patients with separated hemispheres, as performance with both verbal and non-verbal identification is superior in left-nostril stimulation. These results may not necessarily indicate a left-side dominance in odor identification, per se, they could have other possible explanations, such as hemispheric differences in cross-modal comparisons 10 or verbal experimental settings (Gordon, 1974). In addition, odor identification studies in patient groups with focal neuropathology or lobotomy have not shown side-related differences (Carroll, Richardson & Thompson, 1993; Eskenazi, Cain, Novelly & Mattson, 1986). Somewhat unexpected in relation to that, Herz and colleagues reported that odorants presented to the left nostril are more frequently named correctly than odorants presented to the right nostril in healthy participants (Herz, McCall & Cahill, 1999). Neural activity Using the ERP technique, Kobal, Hummel and van Toller (1992) found an interaction between stimulated nostril and odorant type on N1 and P2 latencies. The odorant vanillin elicited longer latencies at left- compared to right-nostril stimulation, whereas the odorant hydrogen sulfide elicited shorter latencies at left- compared to right-nostril stimulation. Since all participants experienced the odor of vanillin as pleasant and the odor of hydrogen sulfide as unpleasant this effect was interpreted as possibly caused by their difference in odor hedonics. A similar result pattern in ERP recordings between stimulated nostrils and odor hedonicity has repeatedly been described by Kobal and colleagues. Thus, they reported larger amplitudes and longer latencies in left- compared to right-nostril stimulation for pleasantly perceived stimuli, and the reversed for unpleasantly perceived stimuli (Kobal, Hummel & Pauli, 1989; Kobal & Hummel, 1991; Hummel & Kobal 1994). Recently, by using a bimodal odorant, that stimulates both the olfactory and trigeminal system, alternating left- and right-nostril stimulation between participants, Lundström and Hummel (2006) found that women generally had larger amplitudes and longer latencies over left- compared to right-hemisphere sites while men showed the reversed pattern. Unfortunately in this respect, no analyses including side of stimulation were reported. In a study that elegantly combined magnetic source imaging and ERP recordings, the overarching result pattern indicated higher lefthemisphere activity in relation to right-hemisphere activity at leftcompared to right-nostril stimulation and higher left-hemisphere activity in relation to right-hemisphere activity for vanilla compared to hydrogen sulfide (Kettenmann, Hummel, Stefan & Kobal, 1997). In neuroimaging, hemispheric differences were revealed in the classical study by Zatorre and colleagues (1992), showing activity corresponding to the piriform cortex in each hemisphere, in the right 11 orbitofrontal cortex and the left inferior medial frontal lobe for birhinal stimulation. Activation in right orbitofrontal cortex, alternatively stronger activation in right than in left orbitofrontal cortex, are one of the most consistently reported results in imaging studies in olfaction, regardless of imaging technique and experimental set-up (e.g., Jones-Gotman et al., 1993; Small et al., 1997; Sobel et al., 1998; Royet et al., 1999; Yousem et al., 1997; Dade, Jones-Gotman, Zatorre, & Evans, 1998; Plailly et al., 2004). Furthermore, this right-side activation has been shown to be present regardless of stimulated nostril (Savic & Gulyas, 2000). Interestingly, monorhinal and birhinal stimulation seems to elicit approximately the same activity pattern (Zatorre & Jones-Gotman, 2000). The notions of this right-hemisphere predominance may be associated with olfactory tasks that tap on perceptual processing. For example, right orbitofrontal cortex activation was found during both pleasantness and intensity judgments (Zatorre, Jones-Gotman & Rouby, 2000). Moreover, Royet and colleagues (1999, 2001) have consistently showed involvement of right orbitofrontal cortex in studies that examined odor familiarity. However, highly aversive odorants do instead show activations in left orbitofrontal cortex and amygdala (Zald & Pardo, 1997; Gottfried, O'Doherty & Dolan, R.J., 2002). Furthermore, Royet et al. (2000) demonstrated increased left hemisphere activity, e.g., in orbitofrontal cortex and superior frontal gyrus, to emotionally valenced odorants, as well as to emotionally valenced visual and auditory stimuli. Accordingly, conducting hedonic judgments of odors has shown more activation in left orbitofrontal cortex compared to passive smelling of the same stimuli (Royet, Plailly, Delon-Martin, Kareken & Segebarth, 2003). In investigating odor identification, Kareken and associates (Kareken, Mosnik, Doty, Dzemidzic & Hutchins, 2003) showed left inferior frontal activity, possibly reflecting semantic analysis, and Savic and colleagues (Savic, Gulyas, Larsson & Roland, 2000) found activation in left insula in discrimination tasks of odor intensity and odor quality. These findings are in agreement with the results by Royet et al. (1999, 2001, 2003) which suggest that complex semantic tasks, that may be associated with naming, show left-hemisphere activation. RESEARCH OBJECTIVES The four studies in this thesis are all designed to provide results that can contribute to extend the knowledge of neuropsychological aspects of 12 human olfaction. More specifically, the main focus has been on the lateralization of olfactory cognitive functions assessed by behavioral testing and by OERP recordings. The lateralization of olfactory functions assessed by behavioral testing was studied by investigating performance in left- versus right-nostril stimulation in healthy participants. Functions explored for lateralization differences were perceived familiarity, recognition memory, memory experience in terms of remember responses that reflect recollection of a specific episode, and know responses that reflect recollection without contextual information, free identification and effects of previous exposure in free identification (Study I). The next paper (Study II) further investigated lateralization effects in free identification and effects of previous exposure on free identification. This study is motivated by findings in Study I regarding repetition priming and lateralization on odor identification, and in relation to these findings somewhat contradictory reports. The aim was to study whether conflicting findings may complement each other and be explained by a broader perspective. The majority of research studies on olfactory functions heavily depend on good stimulus control, stringent presentation procedures and knowledge about possible effects caused by changes in various stimulus dimensions. Historically, this may, in particular, have concerned research using psychophysical methods. However, nowadays this is also of great importance in methods measuring neural activity. The aim of Study III was therefore to extend the knowledge about how a OERP protocol, also to be used in Study IV, is affected by different stimulus durations. The study investigated effects caused by variations (±50 ms) from the commonly used stimulus duration of 200 ms. In order to also investigate if different stimulus durations affect OERPs in populations where clear and reliable registrations can be more crucial to practice, both a group of healthy elderly participants and a group of young healthy subjects were tested. In addition, age differences were studied. The final paper (Study IV) concerns olfactory lateralization by means of OERP registrations in young healthy participants. The protocol used is based on Study III, and in accordance with the result regarding appropriate stimulus duration. The research questions targeted here were particularly OERP activation related to different sites on the coronal plane (i.e., left hemisphere, midline, right hemisphere) and left- versus right-nostril stimulation, and possible interaction effects of these variables. 13 SUMMARY OF EMPIRICAL STUDIES Study I: Lateralization of Olfactory Cognitive Functions: Effects of Rhinal Side of Stimulation Aim This study investigated the olfactory functions of familiarity, recognition memory and identification with respect to behavioral differences between left- and right-nostril stimulation. Based on behavioral research on lateralization of olfactory functions in healthy participants the hypothesized results were a right-side advantage for more sensory-related functions. Since lateralization of higher cognitive functions rarely have been studied, the expected outcome in this respect was less clear. Methods Forty young, right-handed adults (20 men and 20 women) participated who were screened for loss in odor sensitivity. In an initial encoding phase the participant was monorhinally presented with 24 randomly selected odorous stimuli (out of 48 stimuli, all relatively common odors without trigeminal impact) to be rated with respect to familiarity on a bipolar visual analog scale (VAS) ranging from not familiar to very familiar. In a retrieval phase, 10 min later, the initial 24 odors and 24 new odors were presented. The first task was to report whether the odor had been presented in the previous encoding phase. The response time for this was also measured. Next, the participant rated his/her confidence in the response on a VAS, and if the participant responded that the odor had been present in the encoding phase he/she did also report the recollective experience in terms of remember or know. Where remember, reflected a recollection with specific contextual aspects encountered in the study phase, and know, reflected a recollection without any contextual information. Thereafter, the final task was to freely identify the odor by veridical naming. 14 Results The familiarity ratings were significantly higher at right- compared to left-nostril stimulation. However, no side differences were shown in recognition memory for performance (A´), response criterion (B´´), confidence rating, response time, or memory experience (remember/know). Neither were side differences found in identification, although a repetition effect was shown such that previously presented stimuli were significantly more frequently identified compared to new stimuli. See Table 1. Table 1. Mean values for various measures of olfactory cognitive functions. _________________________________________________________________________ Left nostril Right nostril _________________________________________________________________________ Familiarity rating (proportion) 0.54 0.59 Recognition memory Performance (A´) 0.75 0.76 Response criterion (B´´) - 0.36 - 0.35 Confidence rating (proportion) 0.58 0.59 Response time (geometric means; s) Hit 4.35 4.14 Correct rejection 5.87 5.88 False alarm 5.48 5.73 Miss 6.73 7.13 Memory experience (n) Remember response 5.25 5.05 Know response 3.98 4.15 Identification (n) Old stimuli (encoding & retrieval phase) 2.75 2.55 New stimuli (retrieval phase) 1.98 2.20 _________________________________________________________________________ Conclusions The study showed a right-nostril advantage for perceived odor familiarity, supporting previous notations about a right-side advantage for tasks related to odor perception. No side differences were found in episodic recognition or in identification. However, a significant effect of repetition priming in identification was found. This effect seems to be more pronounced for left- compared to right-nostril stimulation, which raises the question about the nature of lateralization in olfactory priming. 15 Study II: Effects of Repetition Priming and Stimulated Nostril on Odor Identification Aim The notation of a possible left-nostril/hemisphere superiority in odor identification has previously been reported from some studies but not from other. Additionally, Study I indicated a left-nostril advantage on priming effects in relation to cognitive olfactory functions. The present study further investigated these two aspects of olfactory lateralization. The expected results were effects of previous exposure in terms of improved performance. It was further expected that this effect of previous exposure would be more pronounced when tested in the left-nostril, and thus revealing a left-nostril advantage in odor identification explained by previous exposure. Methods Fifteen men and 15 women who all were young adults participated. All participants were healthy, right-handed, reported normal chemosensory function and had normal odor detection sensitivity according to tests. In a first session the participant was monorhinally, alternating left- and right-nostril stimulation, presented with 16 odorants to be identified the odor by veridical naming. In a following session, conducted the next day, the same procedure was used with the exception that the eight odorants presented to the left nostril in the first session were presented to the right nostril, and contrary for the other eight odorants. To prevent serial effects or other effects related to the presentation order, lists of 15 pairs (consisting of List A and List B) of randomized stimuli order were used. The participants were matched into pairs in which one participant was presented with List A in the first session and List B in the second session, while the other participant in the pair started with List B followed by List A. Results The initial analyses showed that the identification rate was higher in the second compared to the first session and for left- compared to right16 nostril stimulation, but no interaction effect. Additional analyses that were conducted in order to closer investigate the effects of session and nostril showed no differences between left- and right-nostril stimulation in either the first or second session. Furthermore, no significant differences were found between right-nostril stimulation in session two compared to the other three conditions. However, identification was found to be superior in left-nostril stimulation in session two compared to both right- and left-nostril stimulation in session one. See Figure 3. Analyses of logarithmic response time in correct identifications showed an effect of session, with faster response times in session two, but no nostril or interaction effects. Figure 3. Differences in number of correct identifications at left- and right-nostril stimulation in Session 1 and 2, tested by paired t-tests. Conclusions This study provides further support for the existence of repetition effects on olfactory functions. Noteworthy, these effects seem to be more pronounced when tested in the left nostril. The results do also demonstrate that the left-nostril advantage in odor identification may be explained by effects of repeated testing. 17 Study III: Olfactory Event-Related Potentials from Varying Stimulus Duration in Young and Elderly Adults Aim Stimulus presentation is a factor that has to be carefully considered in order to obtain valid OERP recordings. Several studies have investigated how stimulus dimensions, such as concentration and interstimulus interval, affect the OERPs. However, surprisingly little concern has been shown for stimulus duration, and the issue of temporal summation has been scarcely investigated in the olfactory sense compared to other modalities. One aim of this study was therefore to look closer into how this stimulus parameter affects OERPs in order to evaluate a stimulus protocol for OERP recordings that also would be applied for the investigation of laterality in Study IV. Another aim of the study was to compare different age groups with regards to OERPs, both in order to gain more information about OERP in different age groups per se, and additionally to investigate whether OERP recordings in elderly would benefit from a stimulus protocol with a longer duration. Methods Twenty-one healthy persons participated who were screened for loss in odor sensitivity and constituted two age groups (M=23.7 and M=79.1 years). The odor stimulus consisted of a non-trigeminal concentration of amyl acetate presented by an olfactometer with the stimulus durations of 150, 200 and 250 ms. Neuroelectric activity was recorded at the electrode sites Fz, Cz and Pz (midline). After each stimulus recording the participants rated the perceived odor intensity on the Labeled Magnitude Scale. The participants were presented with the three stimulus durations in blocks separated by a 10 min break. Results The results of a factorial analysis of variance showed no main effect of stimulus duration for latencies of the OERPs, although it showed a significant main effect of age group, such that the elderly had longer latencies than the younger. No significant main effects for duration or age 18 group were revealed by a factorial analysis of variance for OERP amplitudes. Furthermore, there were no significant differences in perceived intensity ratings, neither for duration nor age group, and no interaction between these variables. Conclusions The results suggest that the different stimulus durations used in this study do not have a significant impact on the OERP latencies or amplitudes elicited in healthy young adults. It is further suggested that the typically used duration time of 200 ms in OERPs protocols is appropriate for general purposes including studies of laterality. However, the results indicate that the stimulus duration should perhaps be more carefully considered in OERPs studies of older participants or possibly clinical populations. The study also indicates that these differences in duration time (±50 ms) do not seem to affect the perceived odor intensity. Study IV: Laterality of the Olfactory Event-Related Potential Response Aim This study investigated lateralization in olfaction with the OERP technique. The objectives of the study were twofold, first to better understand whether rhinal side of stimulation has an impact on the latencies and amplitudes of the recorded brain waves. Behavioral studies have demonstrated differences between nostrils in olfactory function, but very few studies have previously focused on differences for rhinal side of stimulation regarding OERPs. The second objective was to study hemispheric differences in odor information processing by means of OERP recordings. Based on results from imaging techniques, in which differences in hemispheric activation are commonly reported in olfactory research, it is of interest to approach this issue also with the OERP method that provide excellent time resolution. 19 Methods Twenty-eight young, right-handed, adults (14 men and 14 women) participated who were screened for loss in odor sensitivity. The participants’ OERPs were recorded for stimulation in both nostrils in separate blocks. The odor stimulus amyl acetate was presented by an olfactometer while neuroelectric activity was recorded at the electrode sites Fz, Cz, Pz (midline), F3, C3, P3 (left hemisphere) and F4, C4, P4 (right hemisphere). Results No significant main effect of rhinal side of stimulation was found on amplitude. An effect of coronal site was revealed, suggesting significantly smaller amplitudes for right-hemisphere sites compared to midline and left-hemisphere sites. Interaction effects were found for peak x nostril and coronal site x peak x nostril. The latter interaction was explained by smaller N1/P2 amplitudes at the right-hemisphere sites in left-nostril stimulation and may explain the first interaction and the effect of coronal site. No significant main effects were shown on latencies, neither for rhinal side of stimulation nor site. Conclusions The present study showed consistent amplitudes and latencies across rhinal sides of stimulation. This finding indicates that stimulus presentations at either the right or the left nostril are sufficient in order for accurate assessment and interpretation of OERPs recordings. Furthermore, some analyses revealed hemispheric differences on amplitude. The results in this perspective are not easily interpreted and imply that more OERPs research in this field is needed for an extended view. 20 GENERAL DISCUSSION The primary aim of this thesis was to investigate lateralization of olfactory cognitive functions and electrophysiological processes. This was conducted by behavioral testing (Study I and Study II) using monorhinal stimulation comparing left- and right-nostril performance. By using the OERP technique, olfactory lateralization was examined by means of stimulated nostril and hemispheric site (Study IV). This was conducted on bases of an evaluated OERP protocol with respect to stimulus duration (Study III). Behavioral findings The olfactory functions targeted in this thesis ranged from perceptually oriented functions in familiarity judgments (Study I) to higher cognitive ones in recognition memory (Study I) and free identification by veridical naming (Study I and Study II). Effects of previous exposure in odor identification were also investigated in both studies. Familiarity ratings were found to be higher at right- compared to left-nostril stimulation. No differences between stimulated nostrils were revealed in episodic recognition memory that was tested with respect to hit performance, response criterion, response confidence and response time. Noteworthy, the reported memory experience showed nominally more know responses in right-nostril stimulation and more remember responses in left-nostril stimulation. Since know responses are associated with perceptual fluency and familiarity-based memory, while remember responses reflect a conceptual, more distinctive, component of recognition, this is in agreement with a right-side predominance of olfactory functions related to perceptual processes. Odor identification and effects of previous exposure in odor identification were investigated in both Study I and II. In Study I no difference was found in identification between left- and right-nostril stimulation. However, an effect of repetition was present, showing that odors presented in both the encoding and retrieval phase were identified at a higher rate compared to odors presented only in the retrieval phase. Subanalyses showed that this priming effect was significant at left-nostril, but not at right-nostril stimulation. To complement these findings in odor identification, this function was measured again in Study II, in two separate sessions with one day in between. Noteworthy, an odorant was 21 in Study I presented to the same nostril in the two sessions and in Study II to the opposite nostril in the two sessions. Study II showed, in accordance to Study I, effects of previous exposure, and did also show effects of rhinal-side of stimulation. Further analyses demonstrated that both these effects are caused by better identification at left-nostril stimulation due to previous exposure. ERP findings Study III examined an OERP protocol with respect to stimulus duration and demonstrated no differences between the investigated durations. Study IV, based on the protocol of Study III, shows consistent amplitudes and latencies regardless of rhinal side of stimulation. Some analyses revealed hemispheric differences in amplitude, suggesting generally higher amplitudes in the parietal site on the left-hemisphere. Importantly, interaction effects were found that indicate that smaller N1/P2 amplitudes at the right-hemisphere sites at left-nostril stimulation can explain the results of side-related differences. Among the findings, these two studies suggest that a standard protocol for OERP recordings using 200 ms stimulus duration can be assessed by either left- or rightnostril stimulation. Lateralization of olfactory function The result in Study I, showing that odors presented to the right nostril are rated as more familiar than odors presented to the left nostril, are in line with several other notions about lateralization differences in human olfaction. This finding agrees well with the notion of a right nostril dominance in other perceptually oriented functions, such as odor discrimination (Zatorre & Jones-Gotman, 1990), pleasantness judgments (Bensafi, Rouby, Farget, Bertrand, Vigouroux & Holley, 2003) and primed edibility judgments (Olsson & Fridén, 2001). The results from imaging studies clearly suggest higher right hemisphere activity related to odor perception (e.g., Small, Jones-Gotman, Zatorre, Petrides & Evans, 1997; Sobel et al., 1998; Yousem et al., 1997; Zatorre et al., 1992). More specifically associated to the present finding, Royet and colleagues (2001) demonstrated higher right orbitofrontal cortex activation in familiarity judgments compared to other olfactory tasks, and Plailly et al. (2005) 22 showed, by contrasting a familiarity task with a detection task, that the odor familiarity judgments activated right piriform cortex. Somewhat related to familiarity and familiarity processing, the finding in Study I of nominally more know responses tested via the right nostril agrees with results from other modalities indication a right hemisphere superiority for processes related to know responses and a left hemisphere superiority for processes related remember responses (Blaxton & Theodore, 1997; Henson, Rugg, Shallice, Josephs & Dolan, 1999). In olfaction, the finding that patients with right-temporal lobe lesions perform more poorly than patients with left-temporal lobe lesions in odor recognition, indicate a right-hemisphere superiority also in odor recognition (Abraham & Mathai, 1983; Jones-Gotman & Zatorre, 1993; Rausch, Serafetinides & Crandall, 1977). Imaging studies point in the same direction; in inspecting odor recognition Savic and colleagues (2000) reported activity in the right piriform cortex but not the left, and Dade et al. (1998) reported unilateral right orbitofrontal activity. However, (a) the general lack of difference between stimulated nostril in memory performance in Study I, (b) the result by Bromley and Doty (1995), that odor recognition memory is better under bilateral than unilateral test conditions, and particularly, (c) the study by Dade, Zatorre and Jones-Gotman (2002), which elegantly combined results in patients with temporal-lobe lesion and imaging results in healthy, indicating an active role of the piriform cortex in odor memory processing and without any clear hemispheric superiority in this function; unitedly suggest that odor recognition memory may normally also involve inter-hemispheric processing. The results of odor identification performance at left- compared to right-nostril stimulation showed no differences between nostrils, neither in Study I or Study II, without previous exposure. These findings are convergent with other findings in healthy participants (Jones-Gotman et al., 1997; Olsson & Cain, 2003), as well as in patients with cerebral lesions (Eskenazi et al., 1986; Carroll, Richardson & Thompson, 1993; Jones-Gotman et al., 1997), and do not support the notion of a leftnostril superiority in odor identification (Herz, McCall & Cahill, 1999; Homewood & Stevenson, 2001). This raises at least two questions. First, why would there be a leftnostril superiority in odor identification? At first sight this seems natural since there is a left-hemisphere specialization in processes related to language (e.g., Cabeza & Nyberg, 2000) and odor identification, at least operationally defined as odor naming taps on these processes. Accordingly, neuroimaging data show left inferior frontal activity during 23 odor identification, possibly reflecting semantic analysis (Kareken et al., 2003) and the results by Royet and colleagues (1999, 2001, 2003) suggest that complex semantic tasks, that may be associated with naming, show left-hemisphere activations. On the other hand, by inspecting the literature about odor identification, the first striking aspect is the poor performance. A classical example is the study by Cain (1979), in which the participants initially freely named only 36 of the 80 presented odors. The approximately same performance, about 50%, in free odor identification is commonly noted in other studies (e.g., Cain, de Wijk, Lulejian, Schiet & See, 1998; Distel & Hudson, 2001, Engen 1987; Larsson & Bäckman, 1997). The difficulty to identify odors have several proposed explanations; one is the notion of a weak linkage between language processes and olfaction (Richardson & Zucco, 1989; Schab, 1991). Interestingly, many studies that have explored this phenomenon indicate difficulties at earlier perceptual stages, such as discrimination and categorization, rather than difficulties in verbalizing or naming the odorant, and perceptual functions in olfaction are commonly noted, if anything, to be predominantly right sided (see Royet & Plailly, 2004). For example, Cain and Potts (1996) showed that failure to identify an odor most probably can depend on a higher cognitive stage involving verbal labeling, as well as on a perceptual stage. Nevertheless, their study suggests that identification difficulties in most cases depend rather on the perceptual failure than on failure associated with the process of verbal labeling. Further support for this view is provided by the paper by Jönsson and colleagues (Jönsson, Tchekhova, Lönner & Olsson 2005) that shows that the inability to correctly name odors is due to failure to recognize the odor rather than to a poor association between the odor and the proper name. Obviously, this does not imply a lack of higher cognitive processes in odor identification, and the task is probably interhemispheric in healthy participants. As a second raised question, why do some studies indicate a leftnostril superiority whereas other studies do not? The conflicting findings can probably be explained by differences in experimental set-ups, as well as in the analyzing and hypothesis testing of the data. In the study by Herz and colleagues (Herz, McCall & Cahill, 1999), that indicate a leftnostril advantage, the hypotheses was tested by a χ2 test, while other studies concerning odor identification commonly use parametric tests with the results analyzed over participants. That study did also include two sessions that permitted effects of previous exposure, something that will be further discussed below. In the study by Homewood and Stevenson (2001), in which 102 participants were tested in large groups, 24 the identification responses were divided into the categories No name provided, Incorrect, Partially correct, and Correct. No differences between the nostrils were found except for the category Correct for which a onetailed t-test reached significance that revealed a left-nostril advantage. An interesting notion to be made is that if the categories Correct and Partially correct are collapsed the mean identification of left- and right-nostril stimulation is exactly the same (see Table 2 in the article). Both Study I and Study II demonstrate effects of repetition priming, where previously exposed odors were identified to a larger extent. Interestingly, this effect is predominantly present when tested by leftnostril stimulation, regardless whether the stimulus initially is presented to the left nostril (Study I) or the right nostril (Study II), demonstrated also by Olsson and Cain (2003). This finding also agree with the notion that left hemisphere is associated with conceptual types of priming in other sensory modalities (Marsolek, 1999). Moreover, based on the effect of previous exposure, the present studies indicate that what at first sight appears to be a left-nostril advantage in odor identification in some cases can be explained by a sequence effect related to the study design. See Figure 4 for illustration. These results additionally display the importance to interpret the results with respect to effects of previous exposure in olfactory studies in general. The findings in study IV demonstrate no main differences for amplitudes or latencies in OERPs with respect to rhinal side of stimulation. This result has recently been showed also in another study including 95 participants (Stuck, Frey, Freiburg, Hörmann, Zahnert & Hummel, 2006). Interestingly, subanalyses in Study IV revealed hemispheric differences on amplitude that suggest generally higher amplitudes in the parietal site on the left-hemisphere. Further inspection indicates that smaller N1/P2 amplitudes at the right-hemisphere sites in left-nostril stimulation can explain these results of side-related differences. A tentative way to explain this finding in relation to lateralization of olfactory function is that left- compared to right-nostril stimulation elicits less activation in the olfactory regions in the right hemisphere known to be particularly involved in perceptual processing. In that case, this finding compliment, and indicate an explanation of the demonstrated rightnostril advantage in tasks related to odor perception. 25 Number of correct identifications 5 4 Left nostril Right nostril 3 2 Broman et al. (2001) 1 0 Latency (log s) of subvocal identifications New odors Old odors 0,5 0,4 0,3 0,2 Olsson & Cain (2003) 0,1 0,0 Control odors Primed odors Number of correct identifications 5 4 3 2 Broman & Wising (submitted) 1 0 Number of correct identifications Session 1 Session 2 ? ? ? ? Herz et al. (1999) Session 1 Session 2 Figure 4. Illustration of studies that have used a design that permits effects of previous exposure when investigating differences in identification between rhinal side of stimulation. 26 Lateralization of olfactory functions has been demonstrated to be related to handedness in some studies (e.g., Hummel, Mohammadian & Kobal, 1998; Youngentob, et al., 1982) but not in others (e.g., Betchen & Doty, 1998; Zatorre & Jones-Gotman, 1990). In a fMRI study by Royet and colleagues (2003), left- and right-handed showed, at least in some areas, reversed activation pattern between hemispheres in response to odors. In general, cognitive functions seem to be less lateralized in lefthanded compared to right-handed individuals (e.g., Laeng & Peters, 1995; Springer & Deutsch, 2003), although the results in olfactory function still are to limited for any conclusions. However, the studies in this thesis cannot compliment this literature, since they only included right handed participants. Olfactory research in general show that when a gender difference is demonstrated women perform better than men (e.g., Cain, 1982; Doty, Applebaum, Zusho & Settle, 1985; Engen, 1987; Larsson, Lövdén & Nilsson, 2003). Regarding gender differences in lateralization of olfactory functions per se, on the basis of the hitherto existing studies, there is no general indication that men and women differ systematically. The present studies that investigate laterality differences (I, II & IV) include an equal number of men and women. Furthermore, all results about laterality differences presented in this thesis have additionally been analyzed with respect to gender without any demonstrated interactions. Conclusions The work in this thesis may contribute to extending the knowledge of psychological functions in human olfaction. The finding that small variations in stimulus duration do not seem to considerably affect OERP waveform is important for the development of a standard OERP protocol. In a similar vein, the included OERP study concerning lateralization showed that a protocol using monorhinal stimulation of either left- or right-nostril probably is sufficient for the majority of applications. That study does also show the usefulness of this method, with satisfying temporal resolution of neural activity, to investigate olfactory lateralization. Although, the obtained results suggest some clarifications, they certainly call for more research in this field with more intricate research questions and more elaborated designs. The finding of a right-nostril advantage in familiarity is in line with results from imaging studies and supports the view of right-side dominance in basic perceptual processing of odors. The idea of a general left-nostril superiority in odor 27 identification without effects of previous exposure is not supported, neither by Study I or by Study II, since none of them initially showed significant effects in identification between nostrils. However, both studies demonstrate consistent effects of previous exposure in odor identification and indicate more pronounced effects tested by the left nostril. 28 REFERENCES Abraham, A. & Mathai, K.V. (1983). The effect of right temporal lobe lesions on matching smells. Neuropsychologia, 21, 277-281. Amoore, J.E. (1963). Stereochemical theory of olfaction. Nature, 198, 271-272. Anderson, J., Maxwell, L. & Murphy, C. (1992). Odorant identification testing in the young child. Chemical Senses, 17, 590. Annett, J.M., Ford, L. & Gifford, M. (1996). Human memory for odour following monorhinal presentation with and without verbal elaboration. Perceptual and Motor Skills, 82, 378. Bensafi, M., Rouby, C., Farget, V., Bertrand, B., Vigouroux, M. & Holley, A. (2003). Perceptual, affective, and cognitive judgments of odors: pleasantness and handedness effects. Brain and Cognition, 51, 270-275. Betchen, S.A. & Doty, R.L. (1998). Bilateral detection thresholds in dextrals and sinistrals reflect the more sensitive side of the nose, which is not lateralized. Chemical Senses, 23, 453-457. Blaxton, T. & Theodore, W.H. (1997). The role of the temporal lobes in recognizing visuospatial material: remembering versus knowing. Brain and Cognition, 35, 5-25. Borg, G. (1982). A category scale with ratio properties for intermodal and interindividual comparisons. In Geissler, H.G. & Petxold, P. (Eds.), Psychophysical Judgement and the Process of Perception. Berlin: VEB Deutxcher Verlag der Wissenschaften. Borg, G. (1998). Borg´s perceived exertion and pain scales. Champaign, IL: Human Kinetics. Borg, E. & Borg, G. (1998). Absolute ME compared with natural CR scaling. Proceeedings of the Fourteenth Annual Meeting of the International Society for Psychophysics, Canada: Universite´ Lavel Québec, 199-204. Bromley, S.M. & Doty, R.L. (1995). Odor recognition memory is better under bilateral than unilateral test conditions. Cortex, 31, 25-40. Buck, L. (2000). The molecular architecture of odor and pheromone sensing in mammals. Cell, 100, 611-618. Buck, L. (2004). The search for odorant receptors. Cell, 116, 117-119. Buck, L. & Axel, R. (1991). A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell, 65, 175-187. 29 Cabeza, R. & Nyberg, L. (2000). Imaging cognition II: An empirical review of 275 PET and fMRI studies. Journal of Cognitive Neuroscience, 12, 1-47. Cain, W.S. (1974). Contribution of the trigeminal nerve to perceived odor magnitude. Annals of the New York Academy of Sciences, 237, 28-34. Cain, W.S. (1979). To know with the nose: keys to odor identification. Science, 203, 467-469. Cain, W.S. (1982). Odor identification by males and females: predictions vs. performance. Chemical Senses, 7, 129-142. Cain, W.S. (1989). Testing olfaction in a clinical setting. Ear, Nose and Throat Journal, 68, 78-86. Cain, W.S. & Gent, J.F. (1991). Olfactory sensitivity: reliability, generality, and association with age. Journal of Experimental Psychology: Human Perception and Performance, 17, 382-391. Cain, W.S. & Potts, B.C. (1996). Switch and bait: probing the discriminative basis of odor identification via recognition memory. Chemical Senses, 21, 35-44. Cain, W.S., de Wijk, R., Lulejian, C., Schiet, F. & See, L.C. (1998). Odor identification: perceptual and semantic dimensions. Chemical Senses, 23, 309-326. Carroll, B., Richardson, J.T.E. & Thompson, P. (1993). Olfactory information processing and temporal lobe epilepsy. Brain and Cognition, 22, 230-243. Cometto-Muñiz, J.E. & Cain, W.S. (1990). Thresholds for odor and nasal pungency. Physiology and Behavior, 48, 719-725. Dade, L.A., Jones-Gotman, M., Zatorre, R.J. & Evans, A.C. (1998). Human brain function during odor encoding and recognition. A PET activation study. Annals of the New York Academy of Sciences, 855, 572-574. Dade, L.A., Zatorre, R.J. & Jones-Gotman, M. (2002). Olfactory learning: convergent findings from lesion and brain imaging studies in humans. Brain, 125, 86-101. Dexter, F. & Chestnut, D.H. (1995). Analysis of statistical tests to compare visual analog scale measurements among groups. Anesthesiology, 82, 896-902. Distel, H. & Hudson, R. (2001). Judgement of odor intensity is influenced by subjects´ knowledge of the odor source. Chemical Senses, 26, 247-251. 30 Doty, R.L. (1975). Intranasal trigeminal detection of chemical vapors by humans. Physiology and Behavior, 14, 855-859. Doty, R.L. (1991). Olfactory system. In Getchell, T.V., Doty, R.L., Bartoshuk, L.M., & Snow, J.B.J. (Eds.), Smell and taste in health and disease. New York: Raven Press. Doty, R.L., Applebaum, S., Zusho, H. & Settle, R.G. (1985). Sex differences in odor identification ability: a cross-cultural analysis. Neuropsychologia, 23, 667-672. Doty, R.L., Bromley, S.M., Moberg, P.J. & Hummel, T. (1997). Laterality in Human Nasal Chemoreception. In Christman, S. (Ed.), Cerebral Asymmetries in Sensory and Perceptual Processing. Amsterdam: North Holland. Doty, R.L., Brugger, W.E., Jurs, P.C., Orndorff, M.A., Snyder, P.J. & Lowry, L.D. (1978). Intranasal trigeminal stimulation from odorous volatiles: psychometric responses from anosmic and normal humans. Physiology and Behavior, 20, 175-185. Doty, R.L., Shaman, P. & Dann, M. (1984). Development of the University of Pennsylvania Smell Identification Test: a standardized microencapsulated test of olfactory function. Physiology and Behavior, 32, 489-502. Engen, T. (1971). Psychophysics: discrimination and detection. In Kling J.W. & Riggs, L.A. (Eds.), Woodworth & Schlosberg´s experimental psychology. New York: Holt, Rinehart and Winston. Engen, T. (1987). Remembering odors and their names. American Scientist, 75, 497-502. Engen, T. (1991). Odor sensation and memory. New York: Praeger Publishers. Eskenazi, B., Cain, W.S., Novelly, R.A. & Mattson, R. (1986). Odor perception in temporal lobe patients with and without temporal lobectomy. Neuropsychologia, 24, 553-562. Evans, W.J., Kobal, G., Lorig, T.S. & Prah, J.D. (1993). Suggestions for collection and reporting of chemosensory (olfactory) event-related potentials. Chemical Senses, 18, 751-756. Fischer, E. & Penzoldt, F. (1886). Über die empfindlichkeit des geruchsinnes. Sitzungsberichte Physikalisch Medizinischen Sozietat Erlangen, 18, 7-10. Gescheider, G.A. (1997). Psychophysics: The Fundamentals. London: Lawrence Erlbaum Associates. 31 Gordon, H. W. (1974). Olfaction and cerebral separation. In Kinsbourne, M. & Smith, W.L. (Eds.), Hemispheric Disconnection and Cerebral Function. Springfield. Gordon, H.W. & Sperry, R.W. (1969). Lateralization of olfactory perception in the surgically separated hemispheres of man. Neuropsychologia, 7, 111-120. Gottfried, J.A., O'Doherty, J. & Dolan, R.J. (2002). Appetitive and Aversive Olfactory Learning in Humans Studied Using EventRelated Functional Magnetic Resonance Imaging, Journal of Neuroscience, 22, 10829-10837. Gottfried, J.A., Winston, J.S. & Dolan, R.J. (2006). Dissociable codes of odor quality and odorant structure in human piriform cortex. Neuron, 49, 467-479. Green, B.G., Dalton, P., Cowart, B., Shaffer, G., Rankin, K. & Higgins, J. (1996). Evaluating the ‘Labeled Magnitude Scale’ for measuring sensations of taste and smell. Chemical Senses, 21, 323-334. Green, B.G., Shaffer, G.S. & Gilmore, M.M. (1993). Derivation and evaluation of a semantic scale of oral sensation magnitude with apparent ratio properties. Chemical Senses, 18, 683-702. Handy, T.C. (2005). Event-Related Potentials -a methods handbook (Ed). London: MIT Press Henning, H. (1916). Der Geruch. Leipzig: Barth. Henson, R.N.A., Rugg, M.D., Shallice, T., Josephs, O. & Dolan, R.J. (1999). Recollection and familiarity in recognition memory: a event-related functional magnetic resonance imaging study. Journal of Neuroscience, 19, 3962-3972. Herz, R.S., McCall, C. & Cahill, L. (1999). Hemispheric lateralization in the processing of odor pleasantness versus odor names. Chemical Senses, 24, 691-695. Homewood, J. & Stevenson, R.J. (2001). Differences in naming accuracy of odors presented to the left and right nostrils. Biological Psychology, 58, 65-73. Hudson, R. (1999). From molecule to mind: the role of experience in shaping olfactory function. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology, 185, 297304. Hummel, T. & Kobal, G. (1994). Chemosensory event-related potentials: effects of dichotomous stimulation with eugenol, and dipydril. In Kurihara K., Suzuki, N. & Ogawa, H. (Eds.), Olfaction and Taste XI. Berlin: Springer Verlag. 32 Hummel, T., Mohammadian, P. & Kobal, G. (1998). Handedness is a determining factor in lateralized olfactory discrimination. Chemical Senses, 23, 541-544. Hummel, T., Sekinger, B., Wolf, S.R., Pauli, E. & Kobal, G. (1997). Sniffin´ sticks: olfactory performance assessed by the combined testing of odor identification, odor discrimination and olfactory thresholds. Chemical Senses, 22, 39-52. Jones-Gotman, M. & Zatorre, R.J. (1993). Odor recognition memory in humans: role of right temporal and orbitofrontal regions. Brain and Cognition, 22, 182-198. Jones-Gotman, M., Zatorre, R.J., Cendes, F., Olivier, A., Andermann, F., McMackin, D., Staunton, H., Siegel, A.M. & Wieser, H.G. (1997). Contribution of medial versus lateral temporal-lobe structures to human odour identification. Brain, 120, 1845-1856. Jones-Gotman, M., Zatorre, R.J., Evans, A.C. & Meyer, E. (1993). Functional activation of right hippocampus during an olfactory recognition task. Society for Neurosciene Abstract, 19, 1002. Jönsson, F.U., Tchekhova, A., Lönner, P. & Olsson, M.J. (2005). A metamemory perspective on odor naming and identification. Chemical Senses, 30, 353-365. Kareken, D.A., Mosnik, D.M., Doty, R.L., Dzemidzic, M. & Hutchins, G.D. (2003). Functional anatomy of human odor sensation, discrimination, and identification in health and aging. Neuropsychology, 17, 482-495. Keller, A. & Vosshall, L.B. (2004). A psychophysical test of the vibration theory of olfaction. Nature Neuroscience, 7, 337-338. Kettenmann, B., Hummel, C., Stefan, H. & Kobal, G. (1997). Multiple olfactory activity in the human neocortex identified by magnetic source imaging. Chemical Senses, 22, 493-502. Kobal, G. (1981). Electrophysiologishe Untersuchungen des Menschlichen Geruchssinns. Stuttgart: Thieme Verlag. Kobal, G. (2003). Electrophysiological measurement of olfactory function. In Doty, R.L. (Ed.), Handbook of Olfaction and Gustation. New York: Marcel Dekker, Inc. Kobal, G. & Hummel, T (1991). Olfactory evoked potentials in humans. In Getchell, T.V., Doty, R.L., Bartoshuk, L.M., & Snow, J.B.J. (Eds.), Smell and taste in health and disease. New York: Raven Press. Kobal, G., Hummel, T. & Pauli, E. (1989). Correlates of hedonic estimates in the olfactory evoked potential. Chemical Senses, 14, 718 33 Kobal, G., Hummel, T., & van Toller, S. (1992). Differences in human chemosensory evoked potentials to olfactory and somatosensory chemical stimuli presented to left and right nostrils. Chemical Senses, 17, 233-244. Kobal, G., van Toller, S. & Hummel, T. (1989). Is there directional smelling? Experientia, 45, 130-132. Koelega, H.S. (1979). Olfaction and sensory asymmetry. Chemical Senses, 4, 89-95. Laeng, B. & Peters, M. (1995). Cerebral lateralization for the processing of spatial coordinates and categories in left- and right-handers. Neuropsychologia, 33, 421-439. Laing, D.G. (1983). Natural sniffing gives optimum odor perception for humans. Perception, 12, 99-117. Larsson, M. & Bäckman, L. (1997). Age-related differences in episodic odor recognition: the role of access to specific odor names. Memory, 5, 361-378. Larsson, M., Lövdén, M. & Nilsson, L.G. (2003). Sex differences in recollective experience for olfactory and verbal information. Acta Psychologica, 112, 89-103. Linnaeus, C. (1756). Odores medicamentorum [Wåhlin, A. (1752)]. Amoenitates Academicae, 3, 183-201. Liu, G., Sobering, G., Duyn, J. & Moonen, C. (1993). A functional MRI technique combining principles of echo-shifting with a train of observations (PRESTO). Magnetic Resonance in Medicine, 30, 764-768. Lundström, J.K. & Hummel, T. (2006). Sex-specific hemispheric differences in cortical activation to a bimodal odor. Behavioural Brain Research, 166, 197-203. Marsolek, C.J. (1999). Dissociable neural subsystems underlie abstract and specific object recognition. Psychological Science, 10, 111-118. Martinez, B.A., Cain, W.S., de Wijk, R.A., Spencer, D.D., Novelly, R.A. & Sass, K.J. (1993). Olfactory functioning before and after temporal lobe resection for intractable seizures. Neuropsychology, 7, 351-363. McLean, J.H. & Shipley, M.T. (1992). Neuroanatomical substrates of olfaction. In Serby, M.J. & Chobor, K.L. (Eds.), Science of Olfaction. New York: Spinger-Verlag. 34 Neely, G., Ljunggren, G., Sylven, C. & Borg, G. (1992). Comparison between the Visual Analogue Scale (VAS) and the Category Ratio Scale (CR-10) for the evaluation of leg exertion. International Journal of Sports Medicine, 13 133-136. Nietzsche, F. (1888). Ecce Homo [Le Guérer, A. (2002), Olfaction, Taste and Cognition]. Nordin, S., Brämerson, A., Lidén, E. & Bende, M. (1998). The Scandinavian Odor Identification Test: development, reliability, validity and normative data. Acta Otolaryngol, 118, 226-234. Ogawa, S., Lee, T.M., Kay, A.R., & Tank, D.W. (1990). Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proceedings of the National Academy of Sciences of the United States of America 87, 9868-9872. Olsson, M.J. & Cain, W.S. (2003). Implicit and explicit memory for odors: Hemispheric differences. Memory and Cognition, 31, 44-50. Olsson, M.J. & Fridén, M. (2001). Evidence of odor priming: Edibility judgements are primed differently between the hemispheres. Chemical Senses, 26, 117-123. Pendense, S.G. (1987). Hemispheric asymmetry in olfaction on a category judgement task. Perceptual and Motor Skills, 64, 495-498. Plailly, J., Bensafi, M., Pachot-Clouard, M., Delon-Martin, C., Kareken, D.A., Rouby, C., Segebarth, C. & Royet, J.P. (2005). NeuroImage, 24, 1032-1041. Poellinger, A., Thomas, R., Lio, P., Lee, A., Makris, N., Rosen, B.R. & Kwong, K.K. (2001). Activation and habituation in olfaction- an fMRI Study. NeuroImage, 13, 547-560. Polich, J. (2003). Detection of Change: Event-Related Potential and fMRI Findings (Ed). New York: Kluwer Academic Publishers. Porter, J., Anand, T., Johnson, B., Khan, R.M. & Sobel, N. (2005). Brain mechanisms for extracting spatial information from smell. Neuron, 47, 581-592. Price, J.L. (1990). Olfactory system. In Paxinos, G. (Ed.), The Human Nervous System. San Diego: Academic Press. Radil, T & Wysocki, C.J. (1998). Spatiotemporal masking in pure olfaction. (1998). Annals of the New York Academy of Sciences, 855, 641-644. Raichle, M.E. (1994). Images of mind: studies with modern imaging techniques. Annual Review of Psychology, 45, 333-356. Rausch, R., Serafetinides, E.A. & Crandall, P.H. (1977). Olfactory memory in patients with anterior temporal lobectomy. Cortex, 13, 445-452. 35 Richardson, J.T.E. & Zucco, G.M. (1989). Cognition and olfaction: a review. Psychological Bulletin, 105, 352-360. Royet, J.P. & Plailly, J. (2004). Lateralization of olfactory processes. Chemical Senses, 29, 731-745. Royet, J.P., Hudry, J., Zald, D.H., Godinot, D., Grégoire, M.C., Lavenne, F., Costes, N. & Holley, A. (2001). Functional neuroanatomy of different olfactory judgments. NeuroImage, 13, 506-519. Royet, J.P., Koenig, O., Gregoire, M.C., Cinotti, L., Lavenne, F., Le Bars, D., Costes, N., Vigouroux, M., Farget, V., Sicard, G., Holley, A., Mauguière, F., Comar, D. & Froment, J.C. (1999). Functional anatomy of perceptual and semantic processing for odors. Journal of Cognitive Neuroscience, 11, 94-109. Royet, J.P., Plailly, J., Delon-Martin, C., Kareken, D.A. & Segebarth, C. (2003). fMRI of emotional responses to odors: influence of hedonic valence and judgement, handedness, and gender. NeuroImage, 20, 713-728. Royet, J.P., Zald, D., Versace, R., Costes, N., Lavenne, N., Koenig, O. & Gervais, R. (2000). Emotional Responses to Pleasant and Unpleasant Olfactory, Visual, and Auditory Stimuli: a Positron Emission Tomography Study. Journal of Neuroscience, 20, 77527759. Savic, I. & Berglund, H (2000). Right-nostril dominance in discrimination of unfamiliar, but not familiar, odours. Chemical Senses, 25, 517-523. Savic, I. & Gulyas, B. (2000). PET show that odors are processed both ipsilaterally and contralaterally to the stimulated nostril. Neuroreport, 11, 2861-2866. Savic, I. Gulyas, B., Larsson, M. & Roland, P. (2000). Olfactory functions are mediated by parallel and hierarchical processing. Neuron, 26, 736-745. Schab, F.R. (1991). Odor memory: Taking stock. Psychological Bulletin, 109, 242-251. Shipley, M.T. & Ennis, M. (1996). Functional organization of olfactory system. Journal of Neurobiology, 30, 123-176. Small, D.M., Jones-Gotman, M., Zatorre, R.J., Petrides, M. & Evans, A.C. (1997). Flavor processing: more than the sum of its parts. NeuroReport, 8, 3913-3917. 36 Sobel, N., Prabhakaran, V., Desmond, J.E., Glover, G.H., Goode, R.L., Sullivan, E.V. & Gabrieli, J.D. (1998). Sniffing and smelling: separate subsystems in human olfactory cortex. Nature, 392, 282286. Sobel, N., Prabhakaran, V., Desmond, J.E., Glover, G.H., Goode, R.L., Sullivan, E.V. & Gabrieli, J.D.E. (1998). Sniffing and smelling: separate subsystems in the human olfactory cortex. Nature, 392, 282-286. Sobel, N., Prabhakaran, V., Zhao, Z., Desmond, J.E., Glover, G.H., Sullivan, E.V. & Gabrieli, J.D. (2000). Time course of odorantinduced activation in the human primary olfactory cortex. Journal of Neurophysiology, 83, 537-551. Springer, S.P. & Deutsch, G. (2003). Left brain right brain. New York: Freeman and company. Stevens, S.S. (1957). On the psychophysical law. Psychological Review, 64, 153-181. Stuck, B.A., Frey, S., Freiburg, C., Hörmann, K., Zahnert, T. & Hummel, T. (2006). Chemosensory event-related potentials in relation to side of stimulation, age, sex, and stimulus concentration. Clinical Neurophysiology, 117, 1367-1375. Teghtsoonian, R., Teghtsoonian, M., Berglund, B. & Berglund, U. (1978). Invariance of odor strength with sniff vigor: an olfactory analogue to size constancy. Journal of Experimental Psychology: Human Perception and Performance, 4, 144-152. Toulouse, E. & Vaschide, N. (1899). L’Asymetrie sensorielle olfactive. Comptes Rendus des Séances et Mémoires de la Société de Biologie, 51, 785-787. Turin, L. (1996). A spectroscopic mechanism for primary olfactory reception. Chemical Senses, 21, 773-791. Wilson, D.A. & Stevenson, R.J. (2003). Olfactory perceptual learning: the critical role of memory in odor discrimination. Neuroscience and Behavioral Reviews, 27, 307-328. Yang, Q.X., Darzinski, B.J., Eslinger, P.J. & Smith, M.B. (1997). Multigradient echo with susceptibility inhomogeneity compensation (MGESIC): demonstration of fMRI in the olfactory cortex at 3.0 T. Magnetic Resonance in Medicine, 37, 331-335. Youngentob, S.L., Kurtz, D.B., Leopold, D.A., Mozell, M.M. & Hornung, D.E. (1982). Olfactory sensitivity: Is there laterality? Chemical Senses, 7, 11-21. 37 Yousem, D.M., Williams, S.C.R., Howard, R.O., Andrew, C., Simmons, A., Allin, M., Geckle, R.J., Suskind, D., Bullmore, E.T., Brammer, M.J. & Doty, R.L. (1997). Functional MR imaging during odor stimulation: preliminary data. Neuroradiology, 204, 833-838. Zald, D.H. & Pardo, J.P. (1997). Emotion, olfaction, and the human amygdala: Amygdala activation during aversive olfactory stimulation. Proceedings of the National Academy of Sciences of the United States of America, 94, 4119-4124. Zald, D.H. & Pardo, J.V. (2000). Functional neuroimaging of the olfactory system in humans. International Journal of Psychophysiology, 36, 165-181. Zatorre, R.J. & Jones-Gotman, M. (1990). Right-nostril advantage for discrimination of odor. Perception & Psychophysics, 47, 526-531. Zatorre, R.J. & Jones-Gotman, M. (2000). Functional imaging of the chemical senses. In Toga, A.W. & Mazziotta, J.C. (Eds.), Brain Mapping: The Systems. San Diego: Academic Press. Zatorre, R.J., Jones-Gotman, M. & Rouby, C. (2000). Neural mechanisms involved in odor pleasantness and intensity judgments. Neuroreport, 11, 2711-2716. Zatorre, R.J., Jones-Gotman, M., Evans, A.C. & Meyer, E. (1992). Functional localization and lateralization of human olfactory cortex. Nature, 360, 339-340. Zucco, G.M. & Tressoldi, P.E. (1989). Hemispheric differences in odour recognition. Cortex, 25, 607-615. Zwaardemaker, H. (1895). Die physiologie des geruchs. Leipzig: Engelmann. 38