Comparison of Rod/Cone Ratio in Three Species of Shorebirds
advertisement

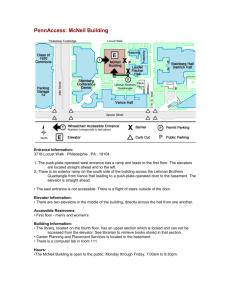
January 1993] Short Communications andCommentaries FORSTER, J.R. 1844. DescriptionesAnimallure quae in itinere ad marls australistetras per annos 1772, 1773 et 1774 susceptocollegit observavitet delineavit JoannesReinoldus Forster (H. Lichtenstein, Ed.). AcademiaLitterarum Regia, Berlin. GMELIN,J.F. 1788-1789. SystemaNaturae, 13th ed. Beer,Lipsiae. HOARE,M.E. 1976. The tactlessphilosopherJohann Reinhold Forster (1729-98). Hawthorn Press, HO^RE,M. E. (Ed.) 1982. The Resolution journal of JohannReinholdForster1772-1775.Hakluyt Society, CambridgeUniv. Press,Cambridge. IREDALE, T. 1938. WilliamAnderson--Ornithologist. 38:60-62. JOHNSON, O. W. 1985. Timing of primary molt in first-yeargolden-ploversand someevolutionary implications.Wilson Bull. 97:237-239. JOHNSON, O. W., ^ND P.M. JOHNSON.1983. Plumagemolt-agerelationshipsin "over-summering"and migratoryLesserGolden-Plovers.Condor85:406419. JOHNSON,O. W., P.M. JOHNSON,AND P. L. BRUNER. 1981. Wintering behavior and site-faithfulness of golden plovers on Oahu. Elepaio 41:123-130. JOHNSON,O. W., M. L. MORTON,P. L. BRUNER,AND P. M. JOHNSON.1989. Fat cyclicity, predicted migratory flight ranges,and featuresof wintering behavior in Pacific Golden-Plovers. Condor 91: 156-177. LANE, B. A. 3 vols.with 2 supplements. Leighand Sotheby, London. LYSAGHT,A. 1959. Some eighteenth century bird paintingsin the library of Sir JosephBanks(17431820). Bull. Br. Mus. Nat. Hist. (Historical Ser.) 1:251-371. MATHEWS,G.M. 1931. John Latham (1740-1837): An early Englishornithologist.Ibis 1 (13th ser.):466475. MEDW^Y, D.G. 1979. Someornithologicalresultsof Melbourne. Emu 141 Cook's third voyage. J. Soc. Bibliog. Nat. Hist. 3:315-351. MEDW^Y, D. G. 1981. The contribution Islands. Pac. Sci. 35:105-175. PENNANT,T. 1785. Arctic zoology. Vol. 2, Birds. Hughs, London (reprinted by Arno Press,1974). SHARPE, R. B. 1906. Birds. Pages79-515 in The history of the collectionscontainedin the natural history departmentsof the British Museum, vol. 2. Trustees of the British Museum, Longmarts, London. STRESEMANN,E. 1949. Birds collected in the North Pacificareaduring Capt.JamesCook'slastvoyage (1778 and 1779). Ibis 91:244-255. STRESEMANN, E. 1950. Birds collectedduring Capt. JamesCook'slastexpedition(1776-1780).Auk 67: 66-88. STRESEMANN, E. 1975. Ornithology from Aristotle to present.Harvard Univ. Press,Cambridge. WInSON, E. 1977. Observations of Hawaiian 1987. Shorebirds in Australia. Nelson, Melbourne. LATHAM,J. 1781-1801. A generalsynopsisof birds. of Cook's third voyageto the ornithologyof the Hawaiian avifauna during Cook'sexpeditions.Elepaio38:13-18. Received 2 October1991,accepted 22 February1992. The Auk 110(1):141-145, 1993 Comparisonof Rod/Cone Ratio in Three Speciesof ShorebirdsHaving Different Nocturnal Foraging Strategies Luz MARINA ROJASDE AZUAJE,•'3 SUSAN T•a,' AND RAYMOND MCNEIL 2 •Institutode Investigaciones en Biomedicina y Ciencias Aplicadas,Universidad de ¸riente, Cumand, Sucre, Venezuela; and 2Dipartementde sciences biologiques, Universit[de Montrial, C. P. 6128, Succ."A", Montreal, QuebecH3C3J7, Canada Nocturnality, the habit of being activeduring darkness,is a characteristicof some aquatic birds, particularly those in the shorebird families (McNeil and Robert 1988, Robert and McNeil 1989, Robert et al. 1989; for detailed review concerning aquatic birds, 3 Present address:D6partement de sciencesbiologiques,Universit• de Montr6al,C. P. 6128,Succ."A", Montr6al, Quebec H3C 3J7, Canada. see McNeil 1991, McNeil et al. 1992, 1993). Two basic types of foraging techniquesare usedby shorebirds: visual searchingfor prey or indicationsof their presence, on or near the surface (e.g. Pluvialisand Charadriusplovers);and probingwith the bill for buried prey that are detectedby tactile and tastecues(e.g. Short-billed Dowitcher, Limnodromus griseus).Some speciesfeed exclusivelywith the sameforagingtechnique both by night and by day (e.g.visualsearching by Pluvialisand Charadrius plovers, or tactile probing 142 Short Communications andCommentaries [Auk,Vol. 110 Milsom1984,Milsometal. 1990),andtheBlack-winged Stilt (McNeil and Robert 1988, Robert and McNeil 1989) take advantageof the moonlight to forage at night (see also McNeil et al. 1992). According to Pienkowski (1982), Grey Plovers (Pluvialissquatarola) have pecking rates lower on very dark moonless nights, comparedwith daylight, but not on moonlit nights.However,ploverstrategists, suchasthe Ringed Plover (Charadrius hiaticula),apparentlyuse sight as the mainmeansof preydetection,evenon dark nights (Pienkowski1983a).Spencer(1953) and Hale (1980) reported that visibility of the moon is not required; Northern Lapwings (except under dense clouds in winter; see Milsom et al. 1990) and Eurasian Curlews 1989).Somespecies,suchasthe SemipalmatedSand- (Numenius arquata)forageon overcastfull-moon nights exactlythe sameason completelyclear nights.Consequently,an importantquestionis:What allowsplover strategiststo foragevisually during darkness? The relatively large eye in relation to head size of plovers,comparedto that of Calidrisspecies,can be regardedasan advantagefor feeding at night (Dugan 1981,Pienkowski1983a,b). Crab-plovers(Dromas)and stone-curlews(Burhinus),which feed at night, and woodcocks(Scolopax), which perform acrobaticnocturnal display flights (see McNeil 1991), also have large eyes. According to Pienkowski (1983b), it is thought that ploversare mainly a low-latitude family piper (Calidrispusilla;Evans 1979), are visual feeders that evolved •' IXXX•XX X7•XXXXX•XXX•XX I I I •11 Ill III I• III ill Fig. 1. Schematicrepresentation of sampling methodologyfor countingrodsand cones. by Short-billed Dowitchers; Evans 1979, Pienkowski 1983a, McNeil and Robert 1988, Robert and McNeil under some conditions and tactile feeders under oth- under arid semidesert conditions where most prey are active only by night. ers (Schneider 1983). Moreover, some shorebirds The retinal visual receptorsof birds, as of all vermodify their foraging strategiesbetween night and tebrates,are rods and cones(Meyer 1977,Tansley and day.Thus, oystercatchers (Haernatopus; Hulscher ! 976, Erichsen1985).Nocturnal birds have a great prepon1982,Sutherland1982)and Tringaspecies(Goss-Cus- derance of rods in their retinas, while up to 80% of tard 1969, 1970, Evans 1979, McNeil and Robert 1988, photoreceptorsin diurnal birds are cones (Tansley Robert and McNeil 1989) are normally sight feeders and Erichsen1985).If ploversare really sightfeeders duringdaylight,but switchto tactileforagingat night. at night, we would expectthem to have proportionDuring daylight and on bright moonlitnights,Black- ately more rods than Haematopus, Hirnantopus, and winged Stilts (Hirnantopus himantopus) usually are viTringa--daylight sight feeders that switch to tactile sual peckers,but usescythelikesweepsof the bill (a foraging at night. The only referenceconcerningthe tactile technique)on moonlessnights or under low rod/cone ratio in shorebirds is that of Rochon-Dumoonlight conditions(McNeil and Robert 1988,Rob- vigneaud (1943). He claimed that rod numbers vary ert and McNeil 1989). with the speciesin diurnal birds, but are high in Comparedto visual"plover strategists," shorebirds Charadrius hiaticulaand Calidrisalpina;both speciesforthat feed by touch (e.g. Limnodrornus) should be relageat night (Pienkowski1983a,Batty1991).The puratively unaffectedby darkness(Dugan 1981, Pien- poseof our studywas to comparethe rod/cone ratio kowski 1981,Goss-Custard 1983).The presenceof nuof the Grey Plover, the Greater Yellowlegs (Tringa merous touch-sensitivenerve endings (e.g. Herbst's rnelanoleuca), and the Short-billed Dowitcher, species corpuscles)at the bill tip (von Bolze 1968,Schwartz- that have different foragingstrategies. kopff 1973,Pienkowski 1983b;seealso Martin 1990), One adult bird of each specieswas collectedin in combinationwith a trigeminal expansionof the coastallagoons(Chacopataand Laguna de Patos)in the state of Sucre, northeastern Venezuela. In the forebrain (Pettigrew and Frost, 1985, Boire 1989), favors touch feeding of many scolopacidspecies(Lirn- field, within a few minutes of collection, the eyes nodrornus,Gallinago,Calidris,etc.). In addition, taste were injected with 2.5% glutaraldehyde in 0.! M (presenceof tastebudsat tip of beak) may play a role phosphatebuffer (pH = 7.2). In order to enhancethe in locating areasrich in prey (Gerritsen et al. 1983, fixation, the eyeswere then punctured at the cornea Van Heezik et al. 1983; see also Martin 1990). and placed in the samefixative for 30 min, the time Predominantly sight feeders such as Pluvialisand neededfor their transportationto the laboratory.FolCharadriusplovers (Pienkowski 1982, 1983a), the lowing immediate enucleation, the retina, still atNorthern Lapwing (Vanellusvanellus;Spencer 1953, tached to the choroidal layer, was cut into nine sec- January 1993] Short Communications andCommentaries tors, using the pecten as a landmark (Fig. 1A), accordingto the procedureof Meyer and May (1973). Each sectorwas subdivided into four portions (Fig. lB) of which only two were retainedfor analysis(Fig. 1C). After 3 h in the fixative, the retinal portions were washedwith phosphatebufferfor 10 min. The total fixation time was lessthan 4 h. Tissueswere postfixed in 1% OsO4in 0.1 M phosphatebuffer for I h, rinsed in phosphatebuffer followed by distilled water, for 10 min each,and dehydrated in grade alcohol series and propylene oxide. The tissueswere successively infiltrated with a 1:1mixture of propylene oxide-epon medium (LX-112) for 12 h, and pure epon medium for another 2 h. Finally, they were embeddedin sil- iconerubbermoldsfilled with epoxiteresin(Fig. 1D) and polymerized in an oven (60øC)for 48 h. TABLE1. Mean rod and cone densities (œ/100 ttm+ SE, n = 45) and rod/cone ratios corresponding to different Retinal sectors retinal and stained with toluidin blue. For each retinal portion, 12 sections (one every 30-40 sections) were mounted on glassslides(Fig. 1E),resulting in 24 sections for each of the nine sectors. Sections were ex- aminedwith a Zeissphotomicroscope. Rodsand cones were countedin one or two fields,238 ttm wide (Fig. 1F), for a total of 45 countsper sector,or 405 per eye. 1975). The mean rod and cone densities and rod/cone ratios correspondingto each different retinal sector are given in Table 1. Rod and cone densities differ significantly between species(rods, F, = 11.031,df = 2 and 24, P < 0.001; cones, F, = 7.90, df = 2 and 24, P < 0.01). As indicated by the Duncan's a posteriori multiple-range test (P = 0.05), the greatestdensityof rods is found in the Grey Plover, followed by the Short-billedDowitcher,while the greatestdensityof conesis in the GreaterYellowlegs,followed by the Grey Plover. Thus, the Grey Plover has the greatest rod/cone ratio, and the Greater Yellowlegshas the lowest. The density of rods and cones tends to be greaterin someretinal sectorsthan in others,but our Cones Rods: cones Grey Plover 1 2 3 4 5 6 16.9 17.2 18.5 17.1 19.3 17.8 7 8 9 17.0 ñ 1.4 16.3 ñ_ 1.0 16.8 +_ 0.5 1 2 3 4 5 6 7 8 9 12.3 13.2 13.4 13.8 14.1 18.4 11.6 12.2 11.5 + + + ñ ñ ñ 1.3 0.7 0.8 1.1 1.3 1.4 1 2 3 4 5 6 7 8 9 11.3 12.3 13.0 21.9 17.2 15.7 14.1 14.0 14.5 12.5 14.9 18.6 13.2 17.1 17.4 + + ñ ñ ñ ñ 1.5 0.8 0.7 2.2 1.9 1.7 1.4:1.0 1.2:1.0 1.0:1.0 1.3:1.0 1.1:1.0 1.0:1.0 17.0 ñ 1.7 1.0:1.0 17.0 + 1.4 17.1 +_ 0.5 1.0:1.0 1.0:1.0 Greater Yellowlegs Mean rod and cone densities (•/100 ttm+ SE) were calculatedfor each sector.Resultswere analyzed using a one-way analysis of variance (ANOVA) and Duncan's a posteriorimultiple-range test (Nie et al. sectors. Rods Semithin sections of 0.7 ttm thick were obtained with an ultramicrotome(Reitcher-Jung, Ultracut-E) 143 ñ 1.3 ñ 1.3 ñ 1.4 ñ 1.3 _+ 1.6 + 2.2 + 2.6 ñ 2.7 ñ 0.8 Short-billed ñ + + + ñ ñ ñ ñ + 1.0 0.9 1.0 1.0 1.! 1.7 1.1 1.1 1.0 19.6 24.0 22.5 17.3 22.2 18.8 13.4 17.3 15.8 ñ ñ ñ ñ ñ ñ ñ ñ ñ 4.6 1.8 1.4 1.8 1.5 2.3 1.8 1.3 1.5 0.6:1.0 0.6:1.0 0.6:1.0 0.8:1.0 0.6:1.0 1.0:1.0 0.9:1.0 0.7:1.0 0.7:1.0 Dowitcher 11.6 14.0 14.4 15.9 16.0 15.5 13.9 14.1 14.5 ñ ñ ñ ñ ñ ñ ñ ñ ñ 1.4 1.4 1.4 1.2 1.2 1.7 1.0 1.4 1.0 1.0:1.0 0.9:1.0 0.9:1.0 1.4:1.0 1.1:1.0 1.0:1.0 1.0:1.0 1.0:1.0 1.0:1.0 (equivalentto sectors1 to 6 in our study;seeFig. 1) of the Grey Plover (Rojasde Azuaje 1991).According to Meyer (1977),"improvedvisual acuityin birdsmay be correlatedwith the presence"in the retina of areas "housingan extremelydensepopulationof conesand rods."Theseso-calledareasof acutevision "represent circumscribedthickeningsof the retina broughtabout presentobjectivewasnot to analyzethe significance by an increasein the number of visual cells (predomof such a difference. Our startinghypothesiswasverified; the Grey Plo- ver, a diurnaland nocturnalsightfeederwith larger eyes,hasmorerodsand a greaterrod/cone ratio than the GreaterYellowlegs,a daylight sight feeder that switchesto tactileforagingat night. The Short-billed Dowitcher,a tactileforagerbothduringdayandnight, is intermediate. The electron microscope revealed that the outer segmentof rodswas longer in the Grey Plover than in the GreaterYellowlegswith the Short-billedDowitcherbeing intermediate(Rojasde Azuaje1991).Furthermore,theserods,with longeroutersegments,are particularly abundant in some areas of the retina inantly cones)" characteristicallylonger than elsewhere in the retina (see also Walls 1942). The outer segments of the photoreceptors contain the visual pigments that are responsiblefor the absorption of light incident upon the retina (Meyer 1977). If areas of longerconesfavorgoodvisualacuityat high light levels(Meyer 1977),it seemslikely that areasof longer rods would be associatedwith better low-light sensitivity. From these results, it appearsthat the Greater Yellowlegs is lesswell adapted for nocturnal vision than either the Grey Plover or the Short-billed Dowitcher. For this reason,the GreaterYellowlegsmay need to abandonits daytimesight-foragingstrategyfor a tac- 144 ShortCommunications andCommentaries [Auk, Vol. 110 availability of their intertidal invertebrate prey. tile one at night, which may still be quite profitable Pages357-366 in Cyclic phenomena in marine since prey availability may be up to 10 times higher plantsand animals(E. Naylor and R. G. Artnoll, at night (Robertand McNeil 1989,McNeil et al. 1992). Eds.). Pergamon Press,Oxford. The Greater Yellowlegs, thus, uses a tactile sidesweepingstrategythat differsfrom the tactileprobing GERRITSEN, A. F. C., Y. M. VAN HEEZIK, AND C. of the Short-billed Dowitcher but, as for the latter, it SwENIqEN.1983. Chemoreceptionin two further Calidrisspecies(C. maritimaand C. canutus).Neth. does not need to see its prey. J. Zool. 33:485-496. Cone density is highest in the Greater Yellowlegs, J. D. 1969. The winter feeding ecolfollowed by the Grey Plover and the Short-billed GOss-CusTARD, ogy of the RedshankTringatotanus.Ibis 111:338Dowitcher, indicating particularly good visual ad356. aptation in the Greater Yellowlegs, but for daylight J.D. 1970. Feedingdispersionin some only. Both the Greater Yellowlegs and the Grey Plo- Goss-CusTARD, overwintering wading birds. Pages3-35 in Social ver forage by sight during daylight. However, the behaviour in birds and mammals (J. H. Crook, Grey Plover feeds on slow-moving or sessileorganisms (in coastalhabitats on marine worms, small mollusks, crustacea, insects, etc.; in the interior on earthworms, seeds, berries, insects; Bent 1929), while the Ed.). Academic Press, London. GOss-CUSTARD, J.D. 1983. Spatialand seasonalvariations in the food supply of waders Charadrii wintering in the British Isles. Proc. 3rd Nordic Congr. Ornithol. 1981:85-96. Greater Yellowlegs forages on fast-swimming prey on most occasions(water insects, small fishes, crustacea; Bent 1927). Consequently, it appears that a HALE, W. G. 1980. Waders. Collins, London. higher cone density would give the Greater Yellow- HULSCHER,J.B. 1976. Location of cockles (Cardium edule L.) by the Oystercatcher (Haematopusoslegsa goodvisual acuity for foraging on fast-moving tralegusL.) in darknessand daylight. Ardea 64: prey during daylight. 292-310. This study was financed by the Universidad de OrJ.B. 1982. The OystercatcherHaematopus iente, the Fundaci0nGran Mariscalde Ayacucho,and HULSCHER, ostralegus as a predator of the bivalve Macoma the grant no. F-60of CONICIT (ConsejoNacionalde balthica in the dutch Waddensea. Ardea 70:89InvestigacionesCientificasy Tecnologicas, Venezue152. la). The participationof McNeil in this study was financed by the Natural Sciencesand Engineering MARTIN,G. 1990. Birds by night. Poyser,London. Research Council of Canada, the Universit• de MonMcNEIL, R. 1991. Nocturnality in shorebirds.Pages 1098-1104 in Acta XX CongressusInternationalis treal, and the Universidad de Oriente through the collaboration agreement between that university in Venezuela and the Universit• de Montreal. We thank Ornithologici. Christchurch, New Zealand Ornithol. Congr. Trust Board,Wellington. Th•r•se Cabanafor helpful commentson the manu- McNEII•, R., P. DRAPEAU, AND J. D. GOss-CUSTARD. 1992. The occurrenceand adaptive significance script. of nocturnal habits in waterfowl. Biol. Rev. Camb. Philos. Soc.In press. LITERATURE CITED McNEIL, R., P. DRAPEAU, AND R. PIEROTTI. 1993. Nocturnality in colonialwaterbirds:Occurrence, specialadaptations,and suspectedbenefits. Current Ornithol. 10: In press. BATTY,L. 1991. Aspectsof the phenologyof waders (Charadrii) on the Ria Formosa,Portugal.Ph.M. McNEIL, R., AND M. ROBERT. 1988. Nocturnal feedthesis,Univ. Wales, Cardiff, United Kingdom. BENT, A.C. 1927. Life histories of North American ing strategiesof someshorebirdspeciesin a tropshore birds, part 1. U.S. Natl. Mus. Bull. 142. ical environment. Pages 2328-2336 in Acta XIX BENt, A.C. 1929. Life histories of North American Congressus Internationalis Ornithologici (H. shore birds, part 2. U.S. Natl. Mus. Bull. 146. Ouellet, Ed.). Ottawa, Ontario, 1986. Univ. Ottawa Press, Ottawa. Boire, D. 1989. Comparaisonquantitative de l'enc•phale, de sesgrandessubdivisionset de relais MEYER,D.B. 1977. The avian eye and its adaptations. visuels,trijumeauxet acoustiques chez28 esp•ces Pages549-611 in The visual systemin vertebrates, d'oiseaux. Th•se de Ph.D., Univ. Montreal, Monvol. VII/5 (F. Crescitelli, Ed.). Springer Verlag, treal. Berlin. DUGAN,P.J. 1981. The importanceof nocturnalfor- MEYER,D. B., AND H. C. MAY. 1973. The topographical distribution of rods and cones in the adult aging in shorebirds:A consequenceof increased invertebrate prey activity. Pages251-260 in Feedchicken retina. Exp. Eye Res. 17:347-355. ing and survivalstrategiesof estuarineorganisms MILSOM,T.P. 1984. Diurnal behaviourof Lapwings (N. V. Jonesand W. J. Wolff, Eds.). Plenum Press, in relation to moon phase during winter. Bird New York. Study 31:117-120. EvANs,P. R. 1979. Adaptations shown by foraging MILSOM,T. P., J. B. A. ROCHARD,AND S. J. POOLE. 1990. shorebirdsto cyclicvariationsin the activityand Activity patternsof Lapwings Vanellusvanellusin January 1993] ShortCommunications andCommentaries relation to the lunar cycle.Ornis Scand.21:147156. NIE, N. H., C. H. HULL, J. G. JENKINS,K. STEINBRENNER, AND D. H. BENT. 1975. SPSSstatistical package for the social sciences. McGraw-Hill, New York. PETrIGREW, J. D., ANDB. J. FROST.1985. A tactile fovea in the Scolopacidae?Brain Behav. Evol. 26:185195. PIENKOWSKI, M.W. 1981. How foragingploverscope with environmental effects on invertebrate be- haviour and availability. Pages179-182in Feeding and survivalstrategiesof estuarineorganisms (N. V. Jonesand W. J. Wolff, Eds.). Plenum Press, New 145 ROJAS DEAZUAJE,L.M. 1991. Proporci6n de conos y bastonesen la retina de tres especiesde aves limlcolasy la relaci6nentre visi6n y estrategia alimenticia. Tesis de M.Sc., Instituto de Inves- tigacionesen Biomedicina y Ciencias Aplicadas, Univ. Oriente, Cuman•, Venezuela. SCHNEIDER, D.C. 1983. The food and feeding of migratory shorebirds.Oceanus26:38-43. SCHWARTZKOPFF, J. 1973. Mechanoreception.Pages 417-477 in Avian biology,vol. 3 (D. S. Farner and J. R. King, Eds.).AcademicPress,New York. SPENCER, K. G. 1953. The Lapwing in Britain. A. Brown & Sons, London. SUTHERLAND, E.J. 1982. Do oystercatchers selectthe York. most profitable cockles? Anita. Behav. 30:857PIENKOWSKI, M.W. 1982. Diet and energy intake of 861. Grey and Ringedplovers,Pluvialissquatarola and K., ANDJ. T. ERICHSEN.1985. Vision. Pages Charadriushiaticula,in the non-breeding season. TANSLEY, 623-629 in A dictionaryof birds (B.Campbelland J. Zool. (Lond.) 197:511-549. E. Lack, Eds.).Poyser,Calton, United Kingdom. PIENKOWSKI, M.W. 1983a. Changesin the foraging VAN HEEZIK, Y. M., A. F. C. GERRITSEN, AND C. pattern of plovers in relation to environmental factors. Anim. Behav. 31:244-264. SWENNEN.1983. The influence of chemoreception on the foraging behaviour of two speciesof PIENKOWSKI, M.W. 1983b. Surfaceactivity of some sandpiper, Calidrisalbaand Calidrisalpina.Neth. intertidal invertebratesin relation to temperaJ. Sea Res. 17:47-56. ture and foraging behaviour of their shorebird YON BOLZE,G. 1968. Anordnung und Bau der predators.Mar. Ecol. Prog. Ser. 11:141-150. herbstschen K6perchen in Limicolenschn•ibeln ROBERT, M., ANDR. MCNEIL. 1989. Comparativeday im Zusammenhangmit der Nahrungsfindung. and night feedingstrategiesof shorebirdspecies Zool. in a tropical environment. Ibis 131:69-79. ROBERT,M., R. MCNEIL, AND A. LEDUC. 1989. Con- ditions and significanceof night feeding in shorebirdsand other water birds in a tropical lagoon. Auk 101:94-101. ROCHON-DuVlGNEAUD, A. 1943. Lesyeux et la vision Anz. 181:313-343. WALLS, G. L. 1942. The vertebrateeyeand its adaptive radiation. Cranbrook Institute of Science, Bloomfield Hills, Michigan. Received 2 December 1991,accepted 6 September 1992. des vertbbrbs. Masson et Cie, Paris. The Auk 110(1):145-148, 1993 ExtendedParent-Offspring Relationshipsin Greenland White-fronted Geese (Anser alb iœrons ltla virostris) STEPHANIEM. WARREN,1 A.D. FOX,' ALYN WALSH,2 AND PADDY O'SULLIVAN2 •TheWildfowlandWetlandsTrust,Slimbridge, Gloucester GL27BT,UnitedKingdom; and 2National ParksandWildlifeService,WexfordWildfowlReserve, NorthSlob,Wexford,Ireland Continuing associations between parents and fledged offspring are uncommon among most bird species.They are, however, especiallyprevalent in territorial species with cooperative breeding (see Brown 1987).Although cooperativebreeding hasnot been recorded in wildfowl, it has long been recognized that juvenile geeseremain with their parents for at leastpart of their firstyear (Phillips 1916,Eider and Eider 1949,Hochbaum1955).Guiding offspring on migrationto traditionalstagingand winteringsites and teachingoffspringvaluableforagingtechniques and social skills, as well as the locations of safe and productive feeding and roosting sites, are common explanationsfor the prolongedparent-offspringrelationshipsin migratorygeeseand swans. GreenlandWhite-frontedGeese(Anseralbifrons fiavirostris; Dalgetyand Scott1948)breedin WestGreenland between64ø and 72øN,and winter exclusively