ENERGY METABOLISM: THE THREE STAGES The term
advertisement

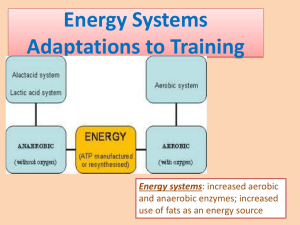
ENERGY METABOLISM: THE THREE STAGES The term metabolism has been used several times in the preceding discussion without completely defining it. It is the process of storing and releasing energy from chemical nutrients. An understanding of energy metabolism is necessary to comprehend the roles that various types of repeat sets, land drills, cycle programs, and other facets of training play in the adaptation process. The two broad categories of metabolism are aerobic and anaerobic. We all presume to understand them; however, they are much more complex than the usual definitions "with oxygen" and "without oxygen" imply. Before we describe aerobic and anaerobic metabolism, we will discuss a third phase - nonaerobic metabolism. Nonaerobic Metabolism This phase of the metabolic process refers to the rapid recycling of ATP from the breakdown of CP. The process has commonly been referred to as anaerobic because it does not require oxygen, but I prefer to use the term "nonaerobic" to differentiate it from the phase of metabolism that involves the anaerobic breakdown of glucose and the formation of lactic acid. The ability to distinguish between these two phases of the metabolic process has important implications for understanding their respective roles in swimming races and training. When a nerve impulse stimulates a muscle fiber to contract, the protein filaments of that fiber-myosin and actin-combine. They activate an enzyme actomyosin ATPase. This enzyme, with water, causes one of the phosphate bonds to be split from the ATP molecule. In the process, the chemical energy in the phosphate bond is released and converted, in part, to mechanical energy that can be used by the muscle fiber to perform the work of contraction. This process is illustrated in Figure 1-3. It is very rapid and does not limit the force that a swimmer can exert. The amount of force is regulated by the number of fibers that are contracting at one time in a muscle. When ATP releases energy for muscular contraction, it also loses one of its phosphate molecules. Consequently, the substance remaining after the breakdown of ATP is termed adenosine diphosphate (ADP) because it now contains only two phosphate molecules (see Figure 1-4). Figure 1-4. The chemical structure of adenosine diphosphate (ADP). The breakdown of one molecule of ATP liberates 7.3 Calories of chemical energy. Some of this is converted to mechanical energy and used by the muscles for contraction, and the rest is converted to heat energy. The percentage of the total energy used for work determines efficiency. For example, when a swimmer's efficiency is listed as 14 percent (a typical efficiency for front crawl stroke swimming, Pendergast et al., 1978), it means that only 14 percent of the chemical energy released is used for the work of muscular contraction. The remaining 86 percent is converted to heat energy. The substance remaining -- ADP -- must be recycled to ATP very rapidly for exercise to continue. Remember, there is only enough ATP in muscle cells to fuel muscular contraction for a few seconds. The recycling of ATP requires replacement of the phosphate and free energy that were lost. The most rapid method for doing this is through the splitting of CP. The phosphate and energy needed for recycling can be made available in one step when this substance is used (see Figure 1-5). ATP can be replaced so rapidly from the breakdown of CP that neither the rate nor the force of muscular contraction will be reduced. The bond that connects creatine and phosphate is broken, so that both energy and phosphate are freed. The phosphate and energy combine with ADP to replace that which was lost so that a new ATP molecule can be formed. After 10 to 15 seconds, when the supply of creatine phosphate is nearly depleted, the processes of anaerobic and aerobic metabolism become the principal sources for ATP replacement. Anaerobic Metabolism This phase of the metabolic process refers to the first eleven steps in the breakdown of glycogen in a process known as glycolysis. The process happens very rapidly and can supply energy for ATP replacement almost as quickly as it is supplied by CP. The steps involved in anaerobic metabolism, as well as the enzymes involved, are listed in Figure 1-6. In most cases, the process begins with the conversion of muscle glycogen to glucose, a procedure that is catalyzed by an activated form of the enzyme phosphorylase. Figure 1-3. The release of energy from ATP. Figure 1-5. The replacement of ATP from creatine phosphate. 1 pH is a measure of the balance between the acidity and alkalinity of fluids. A neutral condition is identified with a value of 7.0. Alkaline fluids have a pH that is greater than this, and fluids that are acidic have a lower pH. We are concerned with the pH of fluids contained within muscle cells (intracellular fluid). When the human body is at rest, the acid-base balance of intracellular fluids is neutral. That is, they have a pH of 7.0. This balance will be upset toward the acidic side when lactic acid is produced during exercise. When muscle pH falls below 7.0, a person is said to be suffering from acidosis. A swimmer's speed will be affected when the fluids within his or her muscles become acidic. The rate of ATP recycling will slow when muscle pH falls below 7.0, with this restraint becoming progressively more severe as it continues to decline. Because it will be impossible for athletes to contract their muscles rapidly and forcefully, their speed will decline. At fast speeds, the accumulation of lactic acid will lower muscle pH well below 7.0 in less than 60 seconds. This is why 100 distances are the upper limit for all-out sprint events. It will take longer to reduce muscle pH when athletes swim longer races at slower speeds. Nevertheless, acidosis will ultimately cause fatigue when the accumulation of lactic acid exceeds its rate of removal from muscles. Figure 1-6. Anaerobic Metabolism After this initial step, the metabolism of glucose proceeds through ten additional stages, ending with the formation of pyruvic acid from phosphophenol-pyruvate. This procedure is catalyzed by the enzyme pyruvate kinase. All of these reactions take place in the protoplasm (cytoplasm) of the muscle cell and do not require oxygen. Another of the important enzymes in this process is phosphofructokinase (PFK), because it plays a major role in regulating the rate of anaerobic metabolism. Hydrogen ions (H+) are also released continuously from glucose when it is metabolized. These are electrically charged atoms that contain energy in the electrons (+) they carry. When pyruvic acid and hydrogen ions combine, they form lactic acid. This reaction is catalyzed by the enzyme lactate dehydrogenase (LDH), particularly the muscle form of that enzyme. Lactic acid is an intermediate product of glucose metabolism and is produced when some of the by-products of that process are not metabolized aerobically. Consequently, it is the end product of the anaerobic phase of metabolism. The accumulation of lactic acid in muscles is thought to be a principal cause of fatigue during swimming races because of its effect on pH. Fortunately, the body can remove some of the lactic acid so that acidosis can be delayed. This happens when lactic acid diffuses out of the muscles and into the blood, where it can be carried to other parts of the body. Some of the lactic acid can even diffuse into fibers within the same muscle group that are not contracting. This process removes lactic acid from working muscle fibers, where the pH is declining, and takes it to other tissues where the pH is normal and additional lactic acid is not being produced. This reduces the rate of fatigue in the working muscles without adversely affecting the other tissues to which the excess lactic acid is taken. Lactic acid is removed from the tissues during recovery by converting it back to pyruvic acid and hydrogen ions. From there, it can be converted back to glycogen and stored for later use or it can enter the aerobic phase of metabolism and be converted to carbon dioxide and water. Another way of delaying fatigue is for the body to produce less lactic acid. All of the pyruvic acid and hydrogen ions released during the anaerobic phase of metabolism do not necessarily have to combine to form lactic acid. The metabolic process can continue in at least two other directions after these two substances are formed and before they combine to make lactic acid. The major direction is through aerobic metabolism. A secondary pathway is through the formation of alanine. Aerobic Metabolism This phase of the metabolic process refers to the remaining steps in glycogen metabolism, ending with the production of carbon dioxide and water. Hundreds of steps are involved in this process. 2 When glycogen is metabolized aerobically, the step producing lactic acid is skipped. Instead, by-products of pyruvic acid and hydrogen ions are metabolized to carbon dioxide and water. Consequently, acidosis is delayed because lactic acid is not produced. Aerobic metabolism has two phases: Krebs cycle (also known as the citric acid cycle) and the electron transport chain. Pyruvic acid is metabolized to carbon dioxide in Krebs cycle. Hydrogen ions are metabolized to water in the electron transport chain. Krebs Cycle Some of the pyruvic acid that was formed during anaerobic metabolism is prepared for entry into Krebs cycle by joining with coenzyme A (A stands for acetic acid) to form acetyl coenzyme A (acetyl-CoA). From there, the acetyl-CoA enters Krebs cycle, where it will ultimately be metabolized to carbon dioxide and water. Within Krebs cycle, acetyl-CoA joins with oxaloacetic acid to form citric acid -the same citric acid found in citrus fruits. The breakdown of citric acid then takes place in a whirling, circular fashion until the acetyl-CoA is dissociated to carbon dioxide and hydrogen ions. The remaining oxaloacetic acid continues through to the starting point, where it picks up more acetyl-CoA to begin the cycle again. A diagram of Krebs cycle is shown in Figure 1-7. Therefore, it is important to remove as many hydrogen ions as possible during exercise so they do not combine with pyruvic acid to form lactic acid and lower muscle pH. A schematic of the electron transport chain is shown in Figure 1-8. In this process, hydrogen ions combine with oxygen to form H2O. The hydrogen ions are prepared for entry into the electron transport chain by combining with another coenzyme — nicotinamide adenine dinucleotide (NAD+) — to form NADH. Still another coenzyme —flavin adenine dinucleotide (FAD) — can also pick up hydrogen ions, forming FADH in the process. Both NADH and FADH are then taken to the electron transport chain, where they are passed through it in bucket brigade fashion by a series of enzymes called the cytochromes. Cytochromes axe composed of iron and protein. The iron (ferric) portion can remove the hydrogen ions from A large number of enzymes regulate Krebs cycle, the most important of which are also shown in Figure 1-7. Endurance training increases the activity of these enzymes so that more pyruvic acid can be taken into Krebs cycle during each minute of exercise so that even less lactic acid is produced at race speeds. The Electron Transport Chain The hydrogen ions that were formed during anaerobic metabolism must also be removed if fatigue is to be delayed. It is the hydrogen ions in lactic acid and not the lactic acid per se that reduces muscle pH. Figure 1-7. Krebs cycle. (Adapted from Costill, 1978, pp. 233-48.) Figure 1-8. The electron transport chain. (Adapted from Lamb, 1978.) 3 NADH and FADH and transfer them to the next cytochrome in the chain. This frees the remaining NAD+ and FAD to return for more hydrogen ions. The energy connected to hydrogen ions is released from the electrons at several points of transfer along the chain and bound to ADP to form ATP. The remaining hydrogen combines with oxygen to form water. Over 90 percent of the recycling of ATP takes place in the electron transport chain. schematic drawing of a muscle fiber, showing the location of mitochondria. It shows the oxygen diffusing into the muscle cell from capillaries, after which it is (transported to the mitochondria by myoglobin. The aerobic endurance of swimmers is in part determined by the size and number of mitochondria they possess. Those swimmers with large and numerous mitochondria will have more little chemical factories for performing aerobic metabolism, so they will not fatigue so quickly. The Role of Oxygen in Aerobic Metabolism Because it is the final acceptor of hydrogen in the electron transport chain, oxygen regulates the rate of energy release from aerobic metabolism. The rate at which NAD+ and FAD can be made available to remove hydrogen ions from muscles depends on the availability of oxygen for releasing the hydrogen molecule from NADH and FADH. When more oxygen is available, a greater amount of hydrogen is dissociated from NADH and FADH, so that more NAD+ and FAD are freed to return and "gather" more hydrogen ions. The Alanine Pathway This is the second pathway that acts to reduce the production of lactic acid. In this process, some of the pyruvic acid and hydrogen ions formed during anaerobic metabolism bind with ammonia to form alanine instead of combining to form lactic acid. The alanine is then transported to the liver, where it can be converted back to glucose and "poured" into the blood to replenish the muscles' energy supply. When oxygen is not available in sufficient quantity to accept all of the hydrogen that was released during exercise, the entire process of aerobic metabolism will back up so that the rates of entry of pyruvic acid into Krebs cycle and of hydrogen ions into the electron transport chain are slowed. When this happens, the hydrogen ions that could not find NAD+ and FAD to combine with will instead combine with excess pyruvic acid to form lactic acid. Unlike anaerobic metabolism, which can proceed in the cytoplasm of muscle cells, the only place where aerobic metabolism can occur is in the mitochondria of those cells. Mitochondria are rod-shaped bodies that are embedded within the cytoplasm (protoplasm) of muscle cells. The mitochondria must have oxygen to perform their function, and that oxygen is transported to them by a protein substance called myoglobin that is in the cytoplasm of muscle cells. Oxygen diffuses through the cell membrane, where it is picked up by myoglobin and transported to the mitochondria. Figure 1-9 is a Fat Metabolism In addition to glycogen, fat and protein can also be metabolized aerobically so that their energy and phosphate can be used to reform ATP. However, these substances must first be converted to an intermediate by-product of glycogen metabolism so that they can enter Krebs cycle. Fats must be converted from triglycerides to free fatty acids for this purpose. Triglycerides are the storage form of fat in the body. They are split to glycerol and three molecules of free fatty acids by the enzyme lipase. The free fatty acids are transported by the blood to working muscles, where they can be absorbed and transported to the mitochondria. They enter the mitochondria with the aid of another enzyme, carnitine transferase (CT). From there, they are converted to acetyl-CoA in a process called beta oxidation. The conversion to acetyl-CoA is catalyzed by the enzyme acetyl-CoA synthetase. Once converted to acetyl-CoA, the free fatty acids are metabolized to carbon dioxide via Krebs cycle in the same way as glycogen. One hundred and forty-seven molecules of ATP can be formed from each fatty acid molecule once it enters Krebs cycle (McArdle, Katch, & Katch, 1981). Protein Metabolism During exercise, certain amino acids-principally alanine are released from working muscles and transported by the blood to the liver, where they are converted to glucose. That glucose is then released into the blood and delivered back to the working muscles. This process is known variously as the alanine-shuttle, or glucose-alanine cycle. Other amino acids that play a similar, though less pronounced, role in supplying energy are glutamate and aspartate. Energy Metabolism During Slow, Medium, and Maximum Swimming Speeds This whole process of metabolism is analogous to filling a tank with fluid until it overflows. Figures l-10a, 1-10b, and l-10c may help clarify this process. Figure 1-9. A muscle fiber, showing the path of oxygen diffusion from capillaries to mitochondria. (Adapted from Edington & Edgerton, Assume that the aerobic phase of metabolism is represented by the large tank and that the smaller overflow container attached to it represents anaerobic metabolism. The breakdown of glycogen and the formation of pyruvic acid and hydrogen ions are represented by the large spigot. The large container has a small spigot at 4 A maximum swimming speed is illustrated in Figure l-10c. In this case, the breakdown of glycogen is so rapid that the capacities of both the large tank and the overflow tank are exceeded. Acidosis causes fatigue rather quickly at this speed because of the rapid accumulation of lactic acid in the overflow tank. That fatigue is apparent because the swimmer is forced to slow his or her speed to one that matches the rate of emptying so that the capacity of the overflow tank is not exceeded. (EN3) The effects of training can also be illustrated by these containers. For example, the time required to fill the large tank depends on the rate fluid is flowing into it versus the rate it is emptying. Training can increase the rate of emptying from the large container by increasing the speed of aerobic metabolism. This, in turn, reduces the amount of lactic acid that pours into the overflow container so that it does not fill as quickly. When this happens, we say that training has reduced the rate of lactic acid accumulation. Training can also increase the rate of emptying from the overflow container so that it does not fill so rapidly. This training effect is stated as an increase in the rate of lactic acid removal from muscles. When these mechanisms are improved, athletes are able to swim faster and longer before acidosis occurs in their muscles. Energy Metabolism Summarized the bottom, but its maximum rate for emptying is slower than the maximum rate at which it can be filled by the larger spigot. The overflow tank also has a small spigot for emptying (into the bloodstream), but its maximum rate of emptying is also slower than the maximum fill rate. At slow speeds, the fluid (pyruvic acid and hydrogen ions) is poured into the large tank only slightly faster than it can be removed, so the tank fills very slowly (see Figure l-10a). This represents easy swimming, where the rate of aerobic metabolism is not maximal, as shown by the fact that the large container is not filled. The excess pyruvic acid and hydrogen ions combine to form a small amount of lactic acid that splashes into the overflow container while the larger container is being filled. However, it can be removed from the overflow container as fast as it pours in, so the athlete should be able to continue at this pace for as long as he or she has glucose, fat, and protein available to supply the energy needed to recycle ATP. (EN1) At medium speeds, the larger (aerobic) container becomes filled and the pyruvic acid and hydrogen ions back up into the overflow container, forming lactic acid (see Figure l-10b). The rate of removal in the anaerobic container nearly matches the rate of accumulation, however, so that it does not fill completely. Consequently, very little lactic acid accumulates in the muscles and acidosis does not occur. This speed represents an overload of aerobic metabolism. It does not represent maximum swimming speed, however. The athlete can still swim faster by using anaerobic metabolism until the smaller container becomes filled. (EN2) Figure 1-11 may make the three phases of metabolism easier to understand. The nonaerobic system is shown at the top, with ATP supplying the energy for muscular contraction, after which it is recycled by CP. The anaerobic system, in the middle, shows the breakdown of glycogen to pyruvic acid. Some of the pyruvic acid combines with hydrogen ions to form lactic acid. Some of that lactic acid remains in the muscles, yet a sizable portion diffuses out of the muscles into the blood, where it is carried to the heart and liver. A portion of the remaining pyruvic acid combines with ammonia to form alanine, and the rest enters Krebs cycle, after first combining with coenzyme A to form acetyl-CoA. The hydrogen ions that remain are also metabolized aerobically. They enter the electron transport chain after combining with NAD+ and FAD to form NADH and FADH. In Krebs cycle, the acetyl-CoA goes through a large number of steps before it is finally converted to carbon dioxide. In the electron transport chain, the hydrogen ions are stripped from NADH and FADH and oxidized to water. This frees additional NAD+ and FAD to return and gather more hydrogen ions. Figure 1-11 illustrates the metabolism of an athlete who is swimming at a sub maximum speed. That speed is, nevertheless, fast enough to overload the aerobic system. This can be seen by the fact that the containers representing both Krebs cycle and the electron transport chain are full. A small amount of lactic acid is being produced in the muscles, as indicated by the shaded area in the container labeled 5 Some experts feel that training at the anaerobic threshold improves aerobic endurance as fast and as much as it can possibly be improved. Furthermore, they believe that training faster may cause athletes to become overtrained. THE EFFICIENCY OF AEROBIC VERSUS ANAEROBIC METABOLISM It is considered more desirable to recycle ATP aerobically than anaerobically for two reasons: (1) because aerobic metabolism results in the replacement of more ATP and (2) because acidosis does not occur during aerobic metabolism. The anaerobic metabolism of one molecule of glycogen produces only four molecules of ATP. The energy from two of these is used to operate the anaerobic process, providing a net of only 2 ATP molecules that can supply energy for muscular contraction. In the aerobic process, one molecule of glycogen provides enough energy to replace 36 molecules of ATP. Anaerobic metabolism produces lactic acid and, consequently, acidosis. On the other hand, the aerobic process produces carbon dioxide and water, which can easily be removed from the body without causing fatigue. Unfortunately, aerobic metabolism cannot supply all of the energy for ATP recycling during swimming races or even during fast training repeats. The process is too slow. Hundreds of steps are needed before that energy can be released. Aerobic metabolism can supplement the energy provided by anaerobic metabolism in these situations. The amount of supplementation will be less in shorter races because more rapid speeds are required, and it will be greater in longer races, in which the pace is somewhat slower. One of the most important functions of training is to increase the rate of aerobic metabolism so that this process can make a greater contribution to the ATP supply in all races. THE INTERACTION OF NONAEROBIC, ANAEROBIC, AND AEROBIC METABOLISM DURING EXERCISE Figure 1-11. The metabolic process. muscles, because the rate at which hydrogen ions are being released is slightly faster than the rate at which they can be transported into the electron transport chain. This small amount of additional lactic acid in the muscles indicates that aerobic metabolism is overloaded. If it were not, the aerobic system would speed up so that the additional hydrogen ions and pyruvic acid could be metabolized aerobically and the formation of additional lactic acid could be avoided. The amount of lactic acid being produced is well within the athlete's capacity, as indicated by the sub maximum level of 3.0 mmol/l of lactic acid in the container labeled muscles and by the fact that the acid-base balance of the muscles is maintained at a neutral pH of 7.0. The circulatory system helps to maintain this low level of lactic acid by diffusing the lactic acid into the bloodstream and carrying it away from the site. When athletes swim at speeds that overload aerobic metabolism without accumulating large amounts of lactic acid, they are said to be training at the anaerobic threshold. Various swimming events are commonly referred to as either aerobic or anaerobic, giving the false impression that the phases of metabolism are separate and that they occur in sequence, with the next phase beginning at the instant the preceding phase has been completed. For example, a common belief is that nonaerobic metabolism supplies all of the energy for muscular contraction during the first 10 to 15 seconds of work. Anaerobic metabolism supposedly takes over for the next 1 or 2 minutes, after which all of the energy is supplied by aerobic metabolism. This is an oversimplification of the actual sequence of events. Actually, all three phases of the metabolic process operate from the first moment of exercise. The only difference is in the relative contribution from each phase. That contribution depends on the speed and the distance of the swim. In sprints, the major contributors are the nonaerobic and anaerobic phases. Aerobic metabolism proceeds too slowly to meet all, or even most, of the rapid demand for energy in these events. Nevertheless, a small amount of the energy for sprinting is supplied by aerobic metabolism. The aerobic contribution becomes greater as the distance increases and/or the athlete swims at a slower speed. Because they are completed at slower average speeds, the major portion of energy for 6 ATP recycling comes from aerobic metabolism in distance races and sub maximum repeat sets in training. Table 1-1. Relative Contributions of Each Phase of Energy Metabolism to Various Swimming Races and Practice Repeats The energy donated by muscle glycogen and by blood glucose and fat during aerobic metabolism is also influenced by distance and swimming speed. During long swims or repeat sets, where the pace is maximum or nearly so (greater than 85 percent of maximum), nearly all of the energy comes from glycogen stored in the working muscle fibers. At slower speeds, both fat and blood glucose provide a greater percentage of the energy. Blood glucose supplies 20 to 30 percent of the energy for endurance training by some estimates (Brooks & Fahey, 1984) and 30 to 40 percent by others (Felig & Wahren, 1975). Fat metabolism probably supplies between 30 to 50 percent of the total energy used during typical two-hour training sessions (Ahlborg, Hagenfeldt, & Wahren, 1974) but only negligible amounts during competition. Only small amounts of energy- 5 to 15 percent-are derived from protein when sufficient muscle glycogen is available (Brooks & Fahey, 1984; Felig & Wahren, 1971). Carbohydrates and fats are the preferred sources of energy for muscular contraction so that protein can be spared to rebuild tissue and enzymes. Therefore, protein becomes a significant source of energy for exercise only when the glucose supply is low. The danger in using excessive amounts of protein for energy lies in the fact that swimmers could literally catabolize their own muscle tissue. They would lose power and endurance if that were to happen. A trained athlete's body always tries to recycle ATP in the most economical manner. That is, it uses aerobic metabolism to the greatest possible extent allowed by the speed of the race or repeat. Within the aerobic process, fat contributes as much energy as the metabolic rate permits. The Contributions of Each Metabolic Phase to Swimming Races and Practice Repeats It is common to refer to sprints as anaerobic events and distance races as aerobic. However, these characterizations are not completely accurate; as explained earlier, all of the different phases of the metabolic process are operating at once in all races. For this reason, it would probably be more accurate to describe the various race distances as mostly aerobic or mostly anaerobic. The values in Table 1-1 estimate the contributions of the different phases of metabolism during competition and training. The aerobic phase of metabolism is subdivided into two phases—glucose metabolism and fat metabolism—to differentiate the roles played by these two substances in supplying energy. Notice that the various races are listed by competition times and distances in the upper part of this table. Sprint and anaerobic repeats are listed by distance and send-off times in the lower section. Also in the lower section, aerobic repeats are listed by set length. The data are presented in this way so that the information can be adapted for swimmers of any age and level of ability. *Neg. = Negligible In the upper portion of the table, the percentage contributions for each energy system that are listed opposite each race distance apply only to experienced senior-level swimmers. They can be misleading when directed at age group and Masters swimmers and to athletes of less than average ability. For swimmers in these categories, competition times are more accurate than distances for reflecting the way their bodies use their various sources of energy. For example, a 10-year-old athlete who swims 100 m in 1:50.00 probably derives energy from the four metabolic phases in approximately the same proportions as a 22-year-old who swims 200 m in the same time. For these reasons, the relationships between race and repeat distances and the contributions from each energy source can only be used as a guide for 7 experienced teenage and senior swimmers. The relationship between swimming time and energy use is more accurate for preadolescents, older Masters swimmers, and competitors with limited experience. Where races are concerned, the nonaerobic and anaerobic phases of metabolism supply most of the energy for events of 25 to 50 yd/m (events requiring 10 to 30 seconds). Anaerobic metabolism is the major contributor for race distances of 100 and 200 yd/m (events lasting 1 to 3 minutes), although the role of aerobic metabolism becomes increasingly more important at the 200 distance. Both anaerobic and aerobic metabolism contribute substantially to the energy supply in races of 400 m and 500 yd (4 to 6 minutes of swimming). The amount of energy supplied non-aerobically and by fat metabolism is negligible at these distances. The aerobic metabolism of glycogen is the chief source of energy for races of 800 meters to 1,650 yards, although anaerobic metabolism contributes one-fourth to one-third of the energy for these distances. The roles of nonaerobic and fat metabolism are, once again, negligible. Where sprint repeat sets are concerned, nonaerobic and anaerobic metabolism supply most of the energy for sprints of 25 yd/m and less. Anaerobic metabolism is the major source during fast swims of 50 and 100 yd/m. The energy for fast repeats of 200 yd/m is obtained almost equally from aerobic and anaerobic sources, with muscle glycogen as the primary source of fuel. Endurance repeat sets can be swum at any distance. The time required to complete the set is the factor that determines the relative contributions from the various sources of energy. Assuming that these sets are swum at the fastest possible average speed—those that require 15 to 20 minutes to complete—a considerable amount of muscle glycogen is used. The energy from this source is derived from both aerobic and anaerobic processes. The contributions from fat (and blood glucose) become significant after the first half hour, when the muscles' glycogen supply is dwindling. After one hour of training, muscle glycogen will be so depleted that fat and blood glucose supply the major portion of the energy for ATP recycling. The energy for speeds that are slow to moderate comes primarily from fat and blood glucose, regardless of the time needed to complete the set. 8