Exam problems
Vous pouvez composer en français.
Veuillez utiliser trois copies séparées, une pour chaque problème. Le premier
problème ne nécessite pas de calculs.
Les notes de cours sont les seuls documents autorisés.
Problem 1 (5 pt)
Bacterial cells can take up the amino acid tryptophan from their surroundings, or, if the external supply is insufficient, they can synthesize tryptophan
from small molecules in the cell. The tryptophan repressor inhibits transcription of the genes in the tryptophan operon, which encodes the tryptophan
biosynthetic enzymes. Upon binding tryptophan, the tryptophan repressor
binds to a site in the promoter of the operon.
a. What would you expect to happen to the regulation of the tryptophan
biosynthetic enzymes in cells that express a mutant form of the tryptophan
repressor that (i) cannot bind to DNA or (ii) binds to DNA even when no
tryptophan is bound to it?
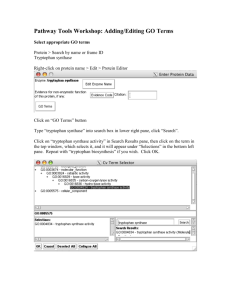
b. In Fig. 1, the bacterial activator protein CAP and the Lac repressor
have been placed in the four possible combinations on their binding sites
in the promoter for the Lac operon. Each combination of gene regulatory
proteins corresponds to a particular mixture of glucose and lactose. For each
of the four combinations, indicate which sugars must be present and whether
the operon is expected to be turned on or off.
c. Imagine that you have created a fusion between the Trp operon, which
encodes the enzymes for tryptophan biosynthesis, and the Lac operon, which
encodes the enzymes necessary for lactose utilization (Fig. 2). Under what
set of conditions (presence or absence of glucose, lactose and tryptophan)
will β-galactosidase be expressed in the strain that carries the fused operon.
Explain your answer. [Hint: Concentrate on the figure].
1
Figure 1: Arrangement of binding sites andthe four possible regulatory proteins on the promoter of the lac operon.
Figure 2: Separated (normal) and fused it Trp and Lac operons.
2
Problem 2 : Reaction-diffusion and optimal spatial distribution of enzyme (7.5 pt)
In some biochemical reactions, enzymes modify substrates in succession. It
can be wondered whether it is better to colocalize enzymes or distribute them
in space to optimize the reaction efficiency. Here, we consider this problem
in a simple one-dimensional setting which could also be relevant for narrow
channels of “lab-on-chip” techniques. Specifically, we consider a substrate S
that is produced at x = 0 by a first enzyme at rate j0 (number of produced
S molecules by second). The molecules diffuse in a one dimensional channel
from x = 0 to x = L and escape irreversibly at x = L (the channel is closed
at x = 0 so that no S escapes toward x < 0) . This effectively acts a sink
at x = L and the concentration is zero there, c(L) = 0. Before S escapes at
x = L it can be transformed into a modified form Sm by a second enzyme
E. We wish to investigate how to distribute a finite quantity of enzyme E
so as to produce the maximum amount of transformed Sm .
1. We first consider the case without enzyme E. The concentration c(x)
of S satisfies the diffusion equation
∂ 2c
∂c
=D 2
∂t
∂x
(1)
1.a. What is the boundary condition obeyed by c at x = 0?
1.b. Which equation is obeyed by c(x) when a stationary state has been
reached?
1.c. Show that the stationary concentration c(x) is given by
c(x) =
j0
(L − x)
D
(2)
2. We now consider the tranformation of S by the enzyme E that we
suppose is distributed with density e(x), so that the total amount of E is
RL
ET = 0 e(x)dx. We write the equation obeyed by S as
∂c
∂ 2c
= D 2 − k e(x) c(x)
∂t
∂x
3
(3)
2.a. Explain what the added last term means. What have we assumed
on the concentration c(x) by writing it in this simple form? What would be
a natural general form?
2.b. When the stationary state has been reached, what is the equation
obeyed by the stationary profile? What is the total rate jm of production of
Sm ? Show that is can be written
jm = j0 + D
∂c
|x=L
∂x
(4)
In the following, we wish to determine the distribution e(x) of the enzyme
E
R L that maximizes jm , for a given total quantity ET of enzyme E, ET =
e(x)dx.
0
3. We consider a distribution in which the concentration e(x) of E is
a uniform step of size L1 : e(x) = e, for 0 ≤ x ≤ L1 , and e(x) = 0, for
L1 ≤ x < L.
3.a. Show that the stationary concentration c(x) is given by a superposition for 0 ≤ x ≤ L1 ,
c(x) = A exp(−x/λ) + B exp(+x/λ)
(5)
√
Show that the diffusion length λ is given by λ = L1 LE with LE = D/(kET ).
What is the relation between A, B and j0 ?
3.b. Write the expression of c(x) for L1 ≤ x ≤ L. What boundary
condition should be satisfied at x = L1 ?
3.c. Determine completely the concentration profile as a function of e
and L1 . Show that the rate of production of jm is given by
#−1
"
r
r
L1
L1
L − L1
sinh(
) + cosh(
)
(6)
jm = j0 1 − √
LE
LE
L1 LE
3.d. We wish to find the enzyme configuration
(i. e.
pthe length L1 ) that
p
maximizes jm . With the notations z = L1 /LE , zL = L/LE , explain why
this is equivalent to finding z, 0 ≤ z ≤ zL that maximizes the function F (z)
with
z2
F (z) = ( L − z) sinh(z) + cosh(z)
(7)
z
4
Show that there are two different cases depending upon whether L is smaller
or larger than L∗ = 3LE .
Note: it may be useful to recall that, for z > 0 tanh(z) > z − z 3 /3.
3.e. Try to qualitatively explain the previous result by comparing the
time to escape from the channel by diffusion to the typical enzymatic modification time for a distributed enzyme configuration.
4. Optional (only in case you have some time left).
4.a. We have found the optimal distribution for a simple enzyme distribution. In order to avoid restricting oneself to particular distribution shapes,
one could try to determine the optimal distribution numerically. Describe
how you would proceed.
4.b. Try to write the general optimisation problem under a form that
could be solved analytically (for instance using Lagrange multipliers). The
solution to this question would have allowed you to write a comment on a
recently published paper, Phys. Rev. Lett. 110, 208104 (2013) (but you will
have to wait for the next opportunity, as others have already done it).
Problem 3 : Proofreading (7.5 pt)
Most chemical reactions in biology are highly specific, involving well-defined
reaction partners. However, unspecific binding can create cross-talk between
reactions and may cause interferences between signaling pathways. Specificity is limited by the equilibrium binding affinity differences between specific
and unspecific partners. However one can improve specificity above equilibrium values using suitable non-equilibrium, energy-consuming schemes. In
this problem, we study an example of such a scheme, called “proofreading”,
in the context of a MAPK cascade, where avoiding cross-talk is an important
requirement.
1. Consider a Michaelis-Menten reaction by which an enzyme KK (e.g.
a MAP kinase kinase) catalyses the phosphorylation of a substrate K (e.g.
a MAP kinase) via the formation of a complex C. The sequence of reactions
with their rates is shown in Fig. 3A. K0 denotes the unphosphorylated version
of K, and K1 its phosphorylation version.
5
A.
B.
Figure 3: Schematic of the phosphorylation reactions. A. K0 gets phosphorylated into K1 by forming a complex with kinase KK. Dephosphorylation
is done by a phosphatase with rate p1 . B. Although K is the main target of
KK, KK can also be involved in the phosphorylation of another substrate
X, which is not its normal target (unspecific activity). The primes 0 denote
the unspecific rates involving X. (Figure from Swain and Siggia, Biophys.
J. 2002.)
1.a. From Fig. 3A (and ignoring part B that involves X), write the
differential equations governing the concentrations of K0 , C0 , C1 , and K1 .
(We drop the brackets [ ] of concentrations for ease of notation.)
1.b. Denote K0∗ = K0 + C0 , K1∗ = K1 + C1 , and Ktot = K0∗ + K1∗ . Show
that at steady state:
C0
K1
f1 K0∗ KK
,
=
b1 + k1 + f1 KK
b3 K1∗
.
=
b3 + f3 KK + p1
(8)
(9)
1.c. Write the differential equation for the concentration K1∗ as a function
of C0 and K1 .
1.d. Explain why the limit b1 k1 can be interpreted as the first reaction being close to equilibrium. In this limit, and further assuming that
phosphatase activity dominates kinase activity, p1 k1 , show that:
K1 ≈ Ktot
k1 f1 KK
.
p1 b1 + f1 KK
6
(10)
2. Now suppose the existence of some competing pathway involving the
phosphorylation of X by the same kinase KK. The reaction are its rates are
shown in Fig. 3B. The binding of X with KK is unspecific compared to that
with K, which is reflected by b01 > b1 , b03 > b3 .
2.a. Call ∆G > 0 the difference of binding free energy between KK and
X0 on the one hand, and KK and K0 on the other hand. Show that:
b1
= exp(−∆G/kB T ),
b01
(11)
where kB is Boltzman’s constant and T is the temperature.
2.b. Write the steady state concentration of X1 as a function of Xtot and
KK.
2.c. We define ρ as the ratio of concentrations of the “error” product
X1 to the “desired” product K1 . Let us assume that both substrates have
the same concentration, Xtot = Ktot . Show that in the limits b1 , b01 k1 ,
p1 k1 discussed above:
ρ≡
X1
b1 + f1 KK
≈ 0
.
K1
b1 + f1 KK
(12)
2.d. Show that the error is lower-bounded by the free energy difference
between the two reactions:
ρ ≥ exp(−∆G/kB T ).
(13)
3. The error ρ can be brought below the free energy limit by having KK
“proofread” K1 through a second reaction, which yields the final product K2
(K1 is then only an intermediate). This scheme is represented in Fig. 4 with
all the rates. The decoy substrate X goes through the same reactions, but
with different off rates b0i > bi (all other rates are the same).
3.a. Redefine K1∗ = K1 + D1 + C1 , K2∗ = C2 + K2 . Show that, when the
rates of the irreversible reactions are much smaller than all others, pi , k1 , w 7
Figure 4: Double phosphorylation. K0 now goes through two phosphorylation steps, to K1 and then to K2 . X goes through equivalent reactions, with
rate b01 , b02 and b03 . (Figure from Swain and Siggia, ibid.)
bi , for all i:
f1 KK
K0∗
b1 + f1 KK
f2 KKb3
=
K∗
b2 f3 KK + b3 b2 + b3 f2 KK 1
b2
D1
=
f2 KK
b3
=
K∗
b3 + f3 KK 2
C0 =
(14)
D1
(15)
K1
K2
(16)
(17)
3.b. The error rate is redefined as ρ = K2 /X2 . In the same limit as above,
where the activity of phosphatases dominates that of kinases, p1 , p2 k1 , w,
show that at steady state:
ρ≈
b1 + f1 KK b2
× .
b01 + f1 KK b02
(18)
(Write the stationary equation for Ki∗ , then use the results of the previous
question to get a system of equations for Ki∗ .)
3.c. How is the fundamental limit of Eq. 13 modified in this case? Where
is energy consumed in this scheme?
8
 0
0