- Wiley Online Library
advertisement

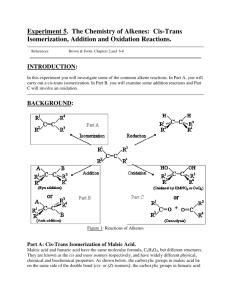
ARTICLE Fumaric Acid Production by Torulopsis glabrata: Engineering the Urea Cycle and the Purine Nucleotide Cycle Xiulai Chen,1,2,3 Jing Wu,1,2,3 Wei Song,1,2,3 Limei Zhang,1,2,3 Hongjiang Wang,1,2,3 Liming Liu1,2,3 1 State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi 214122, China 2 Key Laboratory of Industrial Biotechnology, Ministry of Education, Jiangnan University, Wuxi 214122, China 3 Laboratory of Food Microbial-Manufacturing Engineering, Jiangnan University, Wuxi 214122, China; telephone: þ86-510-85197875; fax: þ86-510-85197875; e-mail: mingll@jiangnan.edu.cn ABSTRACT: A multi-vitamin auxotrophic Torulopsis glabrata strain, a pyruvate producer, was further engineered to produce fumaric acid. Using the genome-scale metabolic model iNX804 of T. glabrata, four fumaric acid biosynthetic pathways, involving the four cytosolic enzymes, argininosuccinate lyase (ASL), adenylosuccinate lyase (ADSL), fumarylacetoacetase (FAA), and fumarase (FUM1), were found. Athough single overexpression of each of the four enzymes in the cytosol improved fumaric acid production, the highest fumaric acid titer (5.62 g L1) was obtained with strain T.G-ASL(H)-ADSL(L) by controlling the strength of ASL at a high level and ADSL at a low level. In order to further improve the production of fumaric acid, the SpMAE1 gene encoding the C4-dicarboxylic acids transporter was overexpressed in strain T.G-ASL(H)-ADSL(L)-SpMAE1 and the final fumaric acid titer increased to 8.83 g L1. This study provides a novel strategy for fumaric acid biosynthesis by utilizing the urea cycle and the purine nucleotide cycle to enhance the bridge between carbon metabolism and nitrogen metabolism. Biotechnol. Bioeng. 2015;112: 156–167. ß 2014 Wiley Periodicals, Inc. Correspondence to: L. Liu Contract grant sponsor: Major State Basic Research Development Program of China Contract grant number: 2013CB733602 Contract grant sponsor: National Natural Science Foundation of China Contract grant number: 31270079 Contract grant sponsor: Provincial outstanding youth foundation of Jiangsu Province Contract grant number: BK2012002 Contract grant sponsor: Program for Innovative Research Team in University Contract grant number: IRT1249 Contract grant sponsor: Fundamental Research Funds for the Central Universities Contract grant number: JUDCF12027 Received 26 April 2014; Revision received 1 June 2014; Accepted 7 July 2014 Accepted manuscript online 24 July 2014; Article first published online 10 October 2014 in Wiley Online Library (http://onlinelibrary.wiley.com/doi/10.1002/bit.25334/abstract). DOI 10.1002/bit.25334 156 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 KEYWORDS: fumaric acid; Torulopsis glabrata; argininosuccinate lyase; adenylosuccinate lyase; genome-scale metabolic model Introduction Fumaric acid, a four-carbon dicarboxylic acid (HO2CCHCHCO2H) that is widely used in the food, pharmaceutical, and chemical industries, is a naturally occurring organic acid (Roa Engel et al., 2008). The U.S. Department of Energy has identified fumaric acid as one of the top 12 biomass building block chemicals that can be converted to high-volume products from renewable sources of carbohydrate (Werpy and Petersen, 2004). Fumaric acid is currently produced on a large scale by microbial fermentation, as this is an environment-friendly, inexpensive, and sustainable development process. Naturally occurring fumaric acid producers, such as Rhizopus formosa, Rhizopus arrhizus, and Rhizopus oryzae, have been isolated, evaluated, and optimized as biocatalysts in the production of fumaric acid by fermentation (Table I). R. formosa has the advantage of requiring low-cost nutrients, but it can produce only 21.3 g L1 fumaric acid (Carta et al., 1999). R. arrhizus can produce 38.0 g L1 fumaric acid with a low yield 0.33 g g1 (Riscaldati et al., 2000). R. oryzae is used as the main producer of fumaric acid and has achieved a high titer and productivity of 56.2 g L1 and 0.7 g L1 h1, respectively (Fu et al., 2010). However, the fermentation process using a rotary reactor proved too complicated for industrial applications (Xu et al., 2012c). Four metabolic engineering strategies have been investigated for producing fumaric acid, and they relate to three ß 2014 Wiley Periodicals, Inc. Table I. Comparison of fumaric acid production by natural and metabolically engineered microorganisms. Strains Fumaric acid (g L1) Yield (g g1) Productivity (g L1 h1) References 21.3 38.0 22.8 37.2 30.2 32.1 41.1 56.2 56.5 40.5 0.34 0.33 0.29 0.53 0.28 0.45 0.48 0.54 0.94 0.51 – 0.46 0.16 1.03 0.19 0.32 0.37 0.70 0.67 0.56 Carta et al. (1999) Riscaldati et al. (2000) Zhou et al. (2014) Zhou et al. (2002) Roa Engel (2010) Kang et al. (2010) Huang et al. (2010) Fu et al. (2010) Wang et al. (2013) Gu et al. (2014) 28.2 25 3.18 1.67 5.64 8.83 0.39 0.78 0.05 0.03 0.11 0.15 0.45 0.26 0.03 0.02 0.06 0.12 Song et al. (2013) Zhang et al. (2012) Xu et al. (2012a) Xu et al. (2012b) Xu et al. (2013a) This study Natural producers R. formosa R. arrhizus R. oryzae Engineered strains E. coli CWF812 R. oryzae ppc S. cerevisiae FMME 001 "PYC2 þ "RoMDH S. cerevisiae FMME 002 DFUM1 þ "RoPYC þ "SFC1 S. cerevisiae FMME 006 DFUM1 þ "RoPYC þ "RoMDH þ "RoFUM1 T. glabrata-ASL(H)-ADSL(L)-SpMAE1 microorganisms, Escherichia coli, R. oryzae, and Saccharomyces cerevisiae (Table I). First, fumaric acid has been produced via the noncyclic glyoxylate route of strategy (i) by combining deletion of the iclR, fumA, fumB and, fumC genes to redirect the carbon flux through the glyoxylate shunt, overexpression of the native ppc gene under the strong tac promoter to enhance the reductive TCA cycle flux and deletion of the arcA and ptsG genes to reinforce the oxidative TCA cycle flux, the final E. coli strain CWF812 allowed production of 28.2 g L1 fumaric acid (Song et al., 2013). Next, strategy (ii) invokes a fumaric acid biosynthetic pathway involving the reductive reactions of the TCA. The carbon flux towards oxaloacetate may be increased through overexpression of endogenous pyruvate carboxylase and exogenous phosphoenolpyruvate carboxylase in R. oryzae. The resultant R. oryzae strain is capable of producing about 25 g L1 fumaric acid (Zhang et al., 2012). In addition, by overexpressing the exogenous RoMDH and RoFUM1 genes and up-regulating the endogenous PYC2 gene, the final engineered S. cerevisiae strain produced 3.18 g L1 fumaric acid (Xu et al., 2012a). Then, strategy (iii) relates fumaric acid production to the oxidation of citrate via the TCA cycle. A final concentration of fumaric acid of 1.67 g L1 was obtained by deletion of the FUM1 gene and overexpression of the RoPYC and SFC1 genes in S. cerevisiae (Xu et al., 2012b). Finally, in strategy (iv), the engineered S. cerevisiae FMME004-6 could produce up to 5.64 g L1 of fumaric acid through simultaneous use of oxidative and reductive routes (Xu et al., 2013a). The multi-vitamin auxotrophic Torulopsis glabrata is a wellestablished microorganism used for the industrial production of pyruvate as it affords three advantages: (i) higher pyruvate production; (ii) higher glucose tolerance; and (iii) higher acid tolerance than S. cerevisiae (Chen et al., 2013). Additionally, the genome-scale metabolic model iNX804 of Candida glabrata (or T. glabrata), consisting of 804 genes, 1287 reactions, and 1025 metabolites, has been reconstructed by genome sequence annotation and biochemical data mining (Xu et al., 2013b). Through model iNX804 a systematical understanding of the physiology and cellular metabolism of T. glabrata may be gained. Furthermore, it has been successfully used to predict the bottleneck of malate production (Chen et al., 2013). In this study, the fumaric acid-related metabolic pathways were identified using model iNX804 and the cytosolic fumaric acid biosynthetic pathways were engineered to enhance fumaric acid production (Fig. 1). The final engineered strain, T.G-ASL(H)-ADSL(L)SpMAE1, was able to produce > 8 g L1 of fumaric acid from glucose. Materials and Methods Strains and Plasmids The multi-vitamin auxotrophic T. glabrata CCTCC M202019 was screened for pyruvate production (Liu et al., 2004b). The engineered yeast strains used in this study (Table II) were derived from T. glabrata CCTCC M202019. T. glabrata CCTCC M202019 Dura3 (T.GDura3) and T. glabrata CCTCC M202019 Dura3Darg8 (T.GDura3Darg8), screened by our laboratory, were used as host strains for gene overexpression (Zhou et al., 2009). E. coli JM109 and shuttle plasmids pYX212, pY26, pY16, and pY2X were used for plasmid construction. Isolation of the Fumarate Biosynthesis Genes The argininosuccinate lyase (ASL, 1386 bp), adenylosuccinate lyase (ADSL, 1449 bp), fumarylacetoacetase (FAA, 786 bp) and fumarase (FUM1, 1452 bp) genes were amplified by PCR using the chromosomal DNA of T. glabrata CCTCC M202019 as template. Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 157 Figure 1. The fumaric acid-related metabolic pathways leading to the formation of fumarate in the cytosol. PYR: pyruvate; OXAL: oxaloacetate; FUM: fumaric acid; MAL: malate; GLU: glutamate; ASP: aspartate; CIT: citrulline; GTP: guanosine triphosphate; ARGSUC: argininosuccinate; ARG: arginine; FUMACAC: fumarylacetoacetate; ACAC: acetoacetate; ADESUC: adenylosuccinate; AMP: adenosine monophosphate; IMP: hypoxanthine nucleotide; TTP Metabolism: tyrosine, tryptophan and phenylalanine metabolism. (A) fumarase; (B) argininosuccinate lyase; (C) fumarylacetoacetase; (D) adenylosuccinate lyase. with the MiniBEST Plasmid Purification Kit Ver. 2.0 (Takara). E. coli JM109 transformations were performed as described by (Inoue et al., 1990). T. glabrata transformations were performed as described by (Zhou et al., 2009). After transformation of the T.GDura3 strain with the plasmids, the yeast strains were plated onto solid medium A. Plasmids pYX212, pYX212(PTPI)-ASL, pYX212(PTPI)-ADSL, pYX212 (PTPI)-FAA, pYX212(PTPI)-FUM1, pY16(PTEF)-ASL, pY26 (PGPD)-ASL, pY26(PTEF)-ASL, pY16(PTEF)-ADSL, pY26 (PGPD)-ASDL, pY26(PTEF)-ASDL, pY16(PTEF)-ASL-(PTEF)ADSL, pY26(PGPD)-ASL-(PTEF)-ADSL, and pY26(PTEF)ASL-(PGPD)-ADSL were transformed into the T.GDura3 strain by electroporation and yielded strains T.G-212, T.G212ASL, T.G-212ADSL, T.G-212FAA, T.G-212FUM1, T.GASL(L), T.G-ASL(M), T.G-ASL(H), T.G-ADSL(L), T.G-ADSL(M), T.G-ADSL(H), T.G-ASL(L)-ADSL(L), T.G-ASL(M)-ADSL(H), T.GASL(H)-ADSL(M), respectively. Furthermore, plasmids pY16 (PTEF)-ASL and pY2X(PGPD)-ADSL, pY16(PTEF)-ASL and pY2X(PTEF)-ADSL, pY2X(PGPD)-ASL and pY16(PTEF)-ADSL, pY26(PGPD)-ASL and pY2X(PGPD)-ADSL, pY2X(PTEF)-ASL and pY16(PTEF)-ADSL, pY26(PTEF)-ASL and pY2X(PTEF)-ADSL, pY2X(PTEF)-ASL-(PGPD)-SpMAE1 and pY16(PTEF)-ADSL, were simultaneously introduced into T.GDura3Darg8, resulting in strains T.G-ASL(L)-ADSL(M), T.G-ASL(L)-ADSL(H), T.G-ASL(M)-ADSL(L), T.G-ASL(M)-ADSL(M), T.G-ASL(H)ADSL(L), T.G-ASL(H)-ADSL(H), T.G-ASL(H)-ADSL(L)SpMAE1, respectively. Media Plasmid Construction and Transformation The plasmids and primers used in this work were given in Tables II and III, respectively. The ASL, ADSL, FAA, and FUM1 genes were amplified from the chromosomal DNA of T. glabrata CCTCC M202019 using the primers BamHI5’ASL&3’ASL-HindIII, EcoRI-5’ADSL&3’ADSL-HindIII, BamHI-5’FAA&3’FAA-HindIII, and BamHI-5’FUM1&3’FUM1-HindIII, respectively. First, the PCR fragments (ASL, FAA, and FUM1 genes) were cut at the introduced BamHI and HindIII sites and ligated to the multicopy plasmid pYX212 that was digested with BamHI and HindIII, resulting in pYX212(P TPI )-ASL, pYX212(P TPI )FAA, and pYX212(P TPI )-FUM1, respectively. In the same way, the plasmids pYX212(P TPI )-ADSL, pY16(P TEF )ASL-(P TEF )-ADSL, pY16(P TEF )-ASL, pY16(P TEF )-ADSL, pY26(P GPD )-ASL, pY26(P TEF )-ASL, pY26(P GPD )-ASDL, pY26(P TEF )-ASDL, pY26(P GPD )-ASL-(P TEF )-ADSL, pY26 (P TEF )-ASL-(P GPD )-ADSL, pY2X(P GPD )-ADSL, pY2XpY2X(P GPD )-ASL, pY2X(P TEF )-ASL, (P TEF )-ADSL, pY2X(P TEF )-ASL-(P GPD )-SpMAE1, were constructed. Restriction endonucleases and the DNA Ligase Kit Ver. 2.0 were used according to the instructions supplied by the manufacturer (Takara). DNA fragments were separated by electrophoresis in a 1% (w/v) agarose gel in Tris-borate/ EDTA. Plasmids were amplified in E. coli JM109 and isolated 158 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 Medium A: the minimal medium (pH 6.0) used for screening contained (per liter) 20 g glucose, 7 g NH4Cl, 5 g KH2PO4, 0.8 g MgSO4-7H2O, 3 g sodium acetate, 32 mg thiamine-HCl, 80 mg biotin, 0.8 mg pyridoxine-HCl, and 16 mg nicotinic acid. Medium B: the medium used for fermentation contained (per liter) 60 g glucose, 7 g NH4Cl, 5 g KH2PO4, 0.8 g MgSO4-7H2O, 6.6 g K2SO4, 3 g sodium acetate, 12 mg thiamine-HCl, 30 mg biotin, 0.4 mg pyridoxine-HCl, and 8 mg nicotinic acid. All vitamins were filtersterilized before addition to the medium. CaCO3 was sterilized by dry-heating sterilization at 160 C for 30 min before use as a pH buffer. Culture Conditions The seed culture inoculated from a slant was cultivated on a reciprocal shaker (200 rpm) at 30 C in a 250 mL flask containing 25 mL medium A for 24 h. The broth was centrifuged, the supernatant liquid was removed and discarded, the pellet was suspended in demineralized water, and the cell suspension was divided equally between flasks containing 25 mL fresh medium B with an initial biomass dry weight of 1 g L1. Fermentation was performed in a 500 mL flask containing 50 mL medium B. The medium was buffered by the addition of 60 g L1 CaCO3 followed by fermentation at 30 C for 72 h with rotation at 200 rpm and all experiments were performed in triplicate. Table II. Torulopsis glabrata strains and plasmids used in this study. Strains and plasmids Strains T.GDura3 T.GDura3Darg8 T.G-212 T.G-212ASL T.G-212ADSL T.G-212FAA T.G-212FUM1 T.G-ASL(L) T.G-ASL(M) T.G-ASL(H) T.G-ADSL(L) T.G-ADSL(M) T.G-ADSL(H) T.G-ASL(L)-ADSL(L) T.G-ASL(L)-ADSL(M) T.G-ASL(L)-ADSL(H) T.G-ASL(M)-ADSL(L) T.G-ASL(M)-ADSL(M) T.G-ASL(M)-ADSL(H) T.G-ASL(H)-ADSL(L) T.G-ASL(H)-ADSL(M) T.G-ASL(H)-ADSL(H) T.G-ASL(H)-ADSL(L)-SpMAE1 Plasmids pYX212 pYX212(PTPI)-ASL pYX212(PTPI)-ADSL pYX212(PTPI)-FAA pYX212(PTPI)-FUM1 pY16 pY16(PTEF)-ASL-(PTEF)-ADSL pY16(PTEF)-ASL pY16(PTEF)-ADSL pY26 pY26(PGPD)-ASL pY26(PTEF)-ASL pY26(PGPD)-ADSL pY26(PTEF)-ADSL pY26(PGPD)-ASL-(PTEF)-ADSL pY26(PTEF)-ASL-(PGPD)-ADSL pY2X pY2X(PGPD)-ADSL pY2X(PTEF)-ADSL pY2X(PGPD)-ASL pY2X(PTEF)-ASL pY2X(PTEF)-SpMAE1 pY2X(PTEF)-ASL-(PGPD)-SpMAE1 Relevant characteristics References CCTCC M202019Dura3 CCTCC M202019Dura3Darg8 CCTCC M202019Dura3 (pYX212) CCTCC M202019Dura3 (pYX212(PTPI)-ASL) CCTCC M202019Dura3 (pYX212(PTPI)-ADSL) CCTCC M202019Dura3 (pYX212(PTPI)-FAA) CCTCC M202019Dura3 (pYX212(PTPI)-FUM1) CCTCC M202019Dura3 (pY16(PTEF)-ASL) CCTCC M202019Dura3 (pY26(PGPD)-ASL) CCTCC M202019Dura3 (pY26(PTEF)-ASL) CCTCC M202019Dura3 (pY16(PTEF)-ADSL) CCTCC M202019Dura3 (pY26(PGPD)-ADSL) CCTCC M202019Dura3 (pY26(PTEF)-ADSL) CCTCC M202019Dura3 (pY16(PTEF)-ASL-(PTEF)-ADSL) CCTCC M202019Dura3Darg8 (pY16(PTEF)-ASL, pY2X(PGPD)-ADSL) CCTCC M202019Dura3Darg8 (pY16(PTEF)-ASL, pY2X(PTEF)-ADSL) CCTCC M202019Dura3Darg8 (pY2X(PGPD)-ASL, pY16(PTEF)-ADSL) CCTCC M202019Dura3Darg8 (pY2X(PGPD)-ASL, pY26(PGPD)-ADSL) CCTCC M202019Dura3 (pY26(PGPD)-ASL-(PTEF)-ADSL) CCTCC M202019Dura3Darg8 (pY2X(PTEF)-ASL, pY16(PTEF)-ADSL) CCTCC M202019Dura3 (pY26(PTEF)-ASL-(PGPD)-ADSL) CCTCC M202019Dura3Darg8 (pY26(PTEF)-ASL, pY2X(PTEF)-ADSL) CCTCC M202019Dura3Darg8 (pY2X(PTEF)-ASL-(PGPD)-SpMAE1, pY16(PTEF)-ADSL) Zhou et al. (2009) Zhou et al. (2009) This study This study This study This study This study This study This study This study This study This study This study This study 2 mm, Amp, URA3, PTPI 2 mm, Amp, URA3, PTPI-ASL 2 mm, Amp, URA3, PTPI-ADSL 2 mm, Amp, URA3, PTPI-FAA 2 mm, Amp, URA3, PTPI-FUM1 CEN6/ARSH4, Amp, URA3, PTEF CEN6/ARSH4, Amp, URA3, PTEF-ASL, PTEF-ADSL CEN6/ARSH4, Amp, URA3, PTEF-ASL CEN6/ARSH4, Amp, URA3, PTEF-ADSL 2 mm, Amp, URA3, PGPD, PTEF 2 mm, Amp, URA3, PGPD-ASL, PTEF 2 mm, Amp, URA3, PGPD, PTEF-ASL 2 mm, Amp, URA3, PGPD-ADSL, PTEF 2 mm, Amp, URA3, PGPD, PTEF-ADSL 2 mm, Amp, URA3, PGPD-ASL, PTEF-ADSL 2 mm, Amp, URA3, PGPD-ASDL, PTEF-ASL 2 mm, Amp, ARG8, PGPD, PTEF 2 mm, Amp, ARG8, PGPD-ADSL, PTEF 2 mm, Amp, ARG8, PGPD, PTEF-ADSL 2 mm, Amp, ARG8, PGPD-ASL, PTEF 2 mm, Amp, ARG8, PGPD, PTEF-ASL 2 mm, Amp, ARG8, PGPD, PTEF-SpMAE1 2 mm, Amp, ARG8, PGPD-SpMAE1, PTEF-ASL Xu et al. (2010) This study This study This study This study Lab collection This study This study This study Xu et al. (2012a) This study This study This study This study This study This study Chen et al. (2013) This study This study This study This study Chen et al. (2013) This study This study This study This study This study This study This study This study This study This study Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 159 Table III. Primers used in this study for gene clone and plasmid construction. Primers Sequence 0 0 5 -CGCGGATCCATGTCAGGTCAAGCTAAAACAAAACTT-30 50 -CCCAAGCTTTTATAACTTCAAGCTCTTTAATTGAGCGAG-30 50 -GGAATTCATGTCTGAGTTTGATAAGTACGCT-30 50 -CCCAAGCTTTCAAACATTTAGTTTAACTTCAGC-30 50 -CGCGGATCCATGAGCTACACTTACCTGAAGGACG-30 50 -CCCAAGCTTCTAAGTTTCTTTGTATTCGTAAGGACC-30 50 -CGCGGATCCATGTTGAGGGCTAGCAGTAGG-30 50 -CCCAAGCTTTTACTTGGTTGGTCCAACCATG-30 50 -AAGGAAAAAAGCGGCCGCATGTCAGGTCAAGCTAAAACAAAACTT-30 50 -TCCCCGCGGTTATAACTTCAAGCTCTTTAATTGAGCGAG-30 50 -AAGGAAAAAAGCGGCCGCATGTCTGAGTTTGATAAGTACGCT-30 50 -TCCCCGCGGTCAAACATTTAGTTTAACTTCAGC-30 BamHI-5 ASL 30 ASL-HindIII EcoRI-50 ADSL 30 ADSL-HindIII BamHI-50 FAA 30 FAA-HindIII BamHI-50 FUM1 30 FUM1-HindIII NotI-50 ASL 30 ASL-SacII NotI-50 ADSL 30 ADSL-SacII Analytical Methods The optical absorbance at 660 nm (A660) was converted to dry cell weight (DCW) according to a predetermined calibration curve (Liu et al., 2004b): A660 : DCW ¼ 1 : 0:23ðgL1 Þ The culture supernatant samples were filtered through 0.22 mm films. Extracellular concentrations of glucose, pyruvate, and fumaric acid were determined by highperformance liquid chromatography (HPLC) as described by (Xu et al., 2012a). The pellets of 10 mL culture broth including cells (OD660 ¼ 9–10) were used to extract intracellular metabolites. AMP was determined using HPLC as described in previous reports (Gusarova et al., 2011). Arginine was determined by HPLC as described by (Xu et al., 2011). GFP Expression Analysis Strains were grown overnight in 25 mL medium B, diluted to OD660 ¼ 0.1 in the same medium and grown for additional hours. The GFP fluorescence intensity of 10,000 yeast cells was measured as described by (Sadeh et al., 2012). cycles, the samples were centrifuged (10,000 g, 15 min, 10 C); the supernatants were collected separately, and the pellets were re-extracted with 2.5 mL precooled 80% (v/v) methanol by vortex for 30 s. After centrifugation, the supernatants were pooled with the first extracts, and the combined extracts were stored at 80 C until further use. Preparation of Cell Extracts and Enzyme Activity Assays Cell extracts were prepared for the determination of enzyme activity (Liu et al., 2004a). ASL activity was determined by the rate of urea synthesis from argininosuccinic acid in the presence of excess arginase as described by (Shih et al., 1969). ADSL activity was determined by measurement of the absorbance at 282 nm (A282) as described by (Lee et al., 1997). FAA activity was determined by hydrolyzing fumarylacetoacetate as described previously (Kvittingen and Brodtkorb, 1986). FUM1 activity produced with L-malate as the substrate was determined by measuring L-malate consumption at 250 nm (A250) as described by (Xu et al., 2012a). Protein concentrations in cell extracts were determined by the Lowry method using bovine serum albumin as the standard (Lowry et al., 1951). Results General Computational Protocol for In silico Prediction of Metabolic Pathways The fumaric acid-related metabolic pathways were searched using the genome-scale metabolic model iNX804 for C. glabrata (or T. glabrata) reconstructed by our laboratory (Xu et al., 2013b). Intracellular Metabolite Extraction Samples were taken with a specialized rapid-sampling setup as described (Canelas et al., 2008). Intracellular metabolites were extracted by freezing-thawing in methanol as described (Canelas et al., 2009). Each sample was re-suspended in 2.5 mL of 80% (v/v) aqueous methanol precooled to 40 C; then, the resulting solution was frozen in liquid nitrogen for 5 min and thawed on ice for 3–5 min. After five freeze-thaw 160 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 Metabolic Pathways for Fumarate Production Ten fumaric acid-related metabolic reactions are contained in T. glabrata GSMM iNX804, which participate in five branches (Table IV): (i) amino acid metabolism; (ii) nucleic acid biosynthesis; (iii) oxidative phosphorylation; (iv) citric acid cycle; and (v) mitochondrial transport reaction. The cytosolic reactions that involve branches (i), (ii), and part of (iii) are catalyzed by FUM1, ASL, ADSL, and FAA (Table IV). FUM1, ASL, and ADSL belong to a superfamily of hydrolases, which share a homotetrameric structure and a similar catalytic mechanism (Trevisson et al., 2009). FUM1 participates in the TCA cycle and catalyzes the reversible dehydration of Lmalate to fumaric acid (Fig. 1A) (Xu et al., 2012c). ASL participates in the urea cycle and catalyzes the reversible breakdown of argininosuccinate to arginine and fumaric acid Arginine and Proline Metabolism Purine and Pyrimidine Biosynthesis Tyrosine, Tryptophan, and Phenylalanine Metabolism Oxidative Phosphorylation Transport, Mitochondrial Transport, Mitochondrial Oxidative Phosphorylation Oxidative Phosphorylation Oxidative Phosphorylation Citric Acid Cycle argsuc[c] < ¼ > arg-L[c] þ fum[c] adesuc[c] < ¼ > amp[c] þ fum[c] 4fumacac[c] þ h2o[c] -> acac[c] þ fum[c] þ h[c] mal-L[c] < ¼ > fum[c] þ h2o[c] fadh2[m] þ fum[c] -> fad[m] þ succ[c] fum[m] þ succ[c] -> fum[c] þ succ[m] fum[m] þ h2o[m] < ¼ > mal-L[m] fadh2[m] þ fum[m] -> fad[m] þ succ[m] fad[m] þ succ[m] < ¼ > fadh2[m] þ fum[m] q6[m] þ succ[m] < ¼ > fum[m] þ q6h2[m] Effect of Single Gene Overexpression on Fumarate Production Abbreviations: c, cytosol; m, mitochondrion. CAGL0I08987g CAGL0B02794g CAGL0J06204g CAGL0A01045g CAGL0I01320g CAGL0M09020g CAGL0A01045g CAGL0I01320g CAGL0L01177g CAGL0C03223g; CAGL0D01958g; CAGL0E03850g; CAGL0F05863g; CAGL0J00847g ASL ADSL FAA FUM1 SYGP SFC1 FUM1 OSM1 SDH SDH argininosuccinate lyase adenylosuccinate lyase fumarylacetoacetase fumarase fumarate reductase, cytosolic/mitochondrial succinate-fumarate transport, mitochondrial fumarase, mitochondrial fumarate reductase succinate dehydrogenase succinate dehydrogenase Subsystem Formula Reactions description GO Gene Table IV. The fumaric acid-related metabolic reactions identified using T. glabrata model iNX804. (Fig. 1B) (Trevisson et al., 2009). ADSL participates in the purine nucleotide cycle and catalyzes two reactions: the conversion of succinylaminoimidazole carboxamide ribotide to aminoimidazole carboxamide ribotide and fumaric acid and the conversion of adenylosuccinate to AMP and fumaric acid (Fig. 1D) (Lee and Colman, 2007). However, FAA belongs to the fumarylacetoacetase family, participates in the catabolism of tyrosine, tryptophan, and phenylalanine in the cytosol and catalyzes the hydrolysis of fumarylacetoacetate to acetoacetate and fumaric acid (Fig. 1C) (Mizutani and Kunishima, 2007). Because of these highlights, ASL, ADSL, and FAA make a bridge between carbon metabolism and nitrogen metabolism, respectively. In addition, FUM1 has been used for fumaric acid production (Xu et al., 2012a; Xu et al., 2012b), whereas ASL, ADSL, and FAA have not. Therefore, the enzymes FUM1, ASL, ADSL, and FAA, which are encoded by CAGL0A01045g, CAGL0I08987g, CAGL0B02794g, and CAGL0J06204g, respectively, were selected for study. Overexpression of the ASL or ADSL gene in the cytosol channeled more pyruvate flux to fumaric acid production. The specific activities of ASL and ADSL in strains T.G212ASL and T.G-212ADSL were 0.124 0.041 U (mg protein)1 and 2.107 0.500 U (mg protein)1, approximately 7-fold and 5-fold higher, respectively, than that of the control strain T.G-212 (Fig. 3). These increases in specific activities were associated with large decreases in the pyruvate titer (Fig. 2A) and significant increases in the fumaric acid titer (Fig. 2B). The final fumaric acid titers were elevated to 0.96 g L1 and 0.61 g L1 in the strains T.G-212ASL and T.G212ADSL, respectively (Fig. 2B). In addition, this single modification in ASL or ADSL in the cytosol improved cell growth and accelerated the glucose consumption rate. DCW was increased by 32.8% and 36.2% for strains T.G-212ASL and T.G-212ADSL, respectively, compared to the control strain T.G-212 (Fig. 2C). The glucose consumption rates for strains T.G-212ASL and T.G-212ADSL were 1.7-fold and 1.3fold higher, respectively, than that of the control strain T.G212 (Fig. 2D). These improvements indicated that the single modification redirected more pyruvate flux to the TCA cycle and improved fumaric acid production through the urea cycle or the purine nucleotide cycle. However, a single overexpression of FUM1 or FAA resulted in an increase in fumaric acid titer (Fig. 2B) and a decrease in pyruvate production (Fig. 2A) that was less than that of either strain T.G-212ASL or T.G-212ADSL, respectively. FUM1 and FAA activities of the strains T.G-212FUM1 and T.G-212FAA increased by about 6.0-fold and 4.5-fold, respectively, compared to the control strain T.G-212 (Fig. 3). The improved activity resulted in minor increases in cell growth (Fig. 2C) and glucose consumption rate (Fig. 2D). All of these slight improvements indicated that the pyruvate flux could not be more effectively redirected to Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 161 Figure 2. The concentrations of metabolites in shake flask cultures of T.G-212 (&), T.G-212FAA (), T.G-212ADSL (D), T.G-212ASL ( ), and T.G-212FUM1 ( ). fumaric acid by overexpressing FUM1 or FAA compared to overexpression of ASL or ADSL, suggesting that further stepwise improvement could be made through overexpression of ASL and ADSL. Figure 3. The specific activity of ASL, ADSL, FAA, and FUM1 in different engineered T. glabrata strains. T.G-212FAA (overexpressing FAA), T.G-212ADSL (overexpressing ADSL), T.G-212ASL (overexpressing ASL), and T.G-212FUM1 (overexpressing FUM1). 162 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 Effect of ASL and ADSL Overexpression on Fumarate Production In order to further investigate the effect of different strengths of a single gene on fumaric acid production, the gene expression strengths were divided into three levels using the GFP reporter, high level (H) under TEF promoter in plasmids pY26 or pY2X, mediate level (M) under GPD promoter in plasmids pY26 or pY2X, and low level (L) under TEF promoter in plasmid pY16 (Fig. 5A). Therefore, the strength of ASL and ADSL were controlled at various levels by different combinations of promoter and plasmid. When the strength of ASL was kept at a high level in all combinations, the concentration of fumaric acid up to 1.23 g L1 was obtained with strain T.G-ASL(H), approximately 28% higher than that of the engineered strain T.G-212ASL (Fig. 4). In addition, the strength of ADSL at a low level improved the fumaric acid titer to 1.82 g L1 with strain T.G-ADSL(L), corresponding to a 4.3-fold increase compared to that of the engineered strain T. G-212ADSL (Fig. 4). These results indicated that fumaric acid production was significantly improved by optimizing the gene expression strengths, suggesting that fumaric acid titers could be further enhanced by simultaneously controlling the ASL and ADSL at different levels. A single modification in ASL or ADSL improved the fumaric acid titer up to 1.23 g L1 or 1.82 g L1, respectively. To further fine-tune fumaric acid synthesis pathway, ASL and Finally, controlling the strength of ASL at a high level and ADSL at a low level obtained a large increase in fumaric acid reaching 5.62 g L1 in the engineered strain T.G-ASL(H)ADSL(L) (Fig. 5C). In addition, this overexpression led to a drastic decrease in the pyruvate titer (Fig. 6), while the DCW of strain T.G-ASL(H)-ADSL(L) corresponded to a 1.14-fold increase compared to that of the control strain T.G-212 and a 16.7% decrease compared to that of strain T.G-ASL(H) (Fig. 6). These results indicated that, upon overexpression of ASL and ADSL, pyruvate flux traveled through the TCA cycle and was effectively channeled to fumaric acid via the urea cycle and the purine nucleotide cycle. Figure 4. The effect of different strengths of a single gene on fumaric acid production. A series of ASL or ADSL expression cassettes were designed at different expression level: high level (H), mediate level (M), and low level (L). ADSL were cytosolically overexpressed in T. glabrata at different expression levels. Based on the above-mentioned division for gene expression levels, a series of ASL and ADSL expression cassettes were designed and assembled in T. glabrata with various strengths to obtain the best metabolic distribution for enhanced fumaric acid production (Fig. 5B). Effect of SpMAE1 Overexpression on Fumarate Production To further improve fumaric acid production, intracellular metabolites were determined for strains T.G-212, T.GASL(H), T.G-ADSL(L), and T.G-ASL(H)-ADSL(L). The concentrations of intracellular fumaric acid, arginine, and AMP in strain T.G-ASL(H)-ADSL(L), were increased by 112.5%, 100.0% and 64.7%, respectively, compared with T.G-212, whereas pyruvate was decreased by 28.0% (Table V). Moreover, these four metabolites in strain T.G-ASL(H)ADSL(L) were also higher than that of the engineered strains T.G-ASL(H) and T.G-ADSL(L). For example, the level of Figure 5. (A) The gene expression was divided into three levels with GFP as reporter: high level (H), mediate level (M), and low level (L). (B) A series of ASL and ADSL expression cassettes were designed at different expression level. (C) The concentrations of fumaric acid were achieved by different ASL and ADSL expression cassettes. Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 163 (Fig. 6). Furthermore, the pyruvate titer was reduced to 3.7 g L1, which was lower than that of strains T.G-ASL(H), T. G-ADSL(L), and T.G-ASL(H)-ADSL(L) (Fig. 6). The concentration of intracellular pyruvate in T.G-ASL(H)-ADSL(L)SpMAE1 was decreased by 25.2% compared with T.GASL(H)-ADSL(L) (Table V). These results indicated that the constructed pathway allowed fumaric acid to be generated out of the pyruvate node and the C4-dicarboxylic acids transporter effectively enabled the export of fumaric acid. Discussion Figure 6. Concentrations of fumaric acid obtained in shake flask cultivation of different strains of T. glabrata overexpressing combinations of ASL, ADSL, and SpMAE1. intracellular fumaric acid in strain T.G-ASL(H)-ADSL(L) was 34.2% and 27.5% higher than that of the engineered strains T. G-ASL(H) and T.G-ADSL(L), respectively (Table V). These increases suggested that the transport capacity of C4dicarboxylic acids probably needed to be enhanced to transport cytosolic fumaric acid out of T. glabrata. The gene SpMAE1 encodes the C4-dicarboxylic acids transporter from S. pombe, which is used to effectively export malate, fumaric acid, and succinate (Zelle et al., 2008). Given this observation, the highest concentration of fumaric acid (up to 8.83 g L1) was obtained with strain T.G-ASL(H)ADSL(L)-SpMAE1 in which the ASL, ADSL and SpMAE1 genes were simultaneously overexpressed (Fig. 6). This dramatic result corresponded to a 67.9-fold increase in the fumaric acid titer compared to that of the control strain T.G-212 and a 1.57-fold increase compared to that of strain T.G-ASL(H)ADSL(L) (Fig. 6). The concentration of intracellular fumaric acid was 0.28 mg/gDCW in T.G-ASL(H)-ADSL(L)-SpMAE1, which was almost concordant with that of the control strain T.G-212 (0.24 mg/gDCW) (Table V). In addition, the DCW of strain T.G-ASL(H)-ADSL(L)-SpMAE1 was reduced by 11.9% compared to that of strain T.G-ASL(H)-ADSL(L), which was similar to that of the control strain T.G-212 In order to effectively produce fumaric acid, all of the fumaric acid biosynthetic pathways were searched using the T. glabrata genome-scale metabolic model iNX804. Among those pathways, the cytosolic biosynthesis pathways were selected to compute the flux distribution and engineered to investigate their influence on fumaric acid production. Finally, the urea cycle and the purine nucleotide cycle were demonstrated to be the most effective pathways for producing fumaric acid. A high fumaric acid titer (5.62 g L1) was obtained with strain T.G-ASL(H)-ADSL(L), in which ASL and ADSL genes were simultaneously overexpressed by controlling the strength of ASL at a high level and ADSL at a low level, respectively. In order to further increase the production of fumaric acid, the SpMAE1 gene was overexpressed in strain T.G-ASL(H)-ADSL(L)-SpMAE1 and the final fumaric acid titer increased to 8.83 g L1. These results lay a good foundation for further study of the fumaric acid metabolic pathways. The most pragmatic systems-based tool for metabolic engineering is the in silico genome-scale metabolic model. This tool has been widely adopted for searching metabolic pathways, modeling cell growth, and predicting flux distribution (Blazeck and Alper, 2010). In this study, T. glabrata model iNX804 was used to search the fumaric acidrelated biosynthetic pathways and revealed 10 fumaric acidrelated metabolic reactions. These reactions may be divided into three categories: cytosolic reactions, mitochondrial reactions, and transport reactions. The cytosolic biosynthesis pathways were selected for further research, due to the fact that they had been successfully used for malate production in T. glabrata (Chen et al., 2013) and utilized to produce fumaric acid in S. cerevisiae (Zelle et al., 2008). A constraint-based genome-scale metabolic model iNX804 was applied to compute the flux distribution in the cytosolic reactions, in Table V. The concentrations of intracellular metabolites in the engineered strains. Strains T.G-212 T.G-ASL(H) T.G-ADSL(L) T.G-ASL(H)-ADSL(L) T.G-ASL(H)-ADSL(L)-SpMAE1 164 Pyruvate (mg/gDCW) Fumaric acid (mg/gDCW) Arginine (mg/gDCW) AMP (mg/gDCW) 7.82 1.04 6.05 1.21 5.92 0.77 6.11 0.74 4.88 0.15 0.24 0.04 0.38 0.01 0.40 0.03 0.51 0.05 0.28 0.02 0.31 0.04 0.55 0.03 0.35 0.04 0.62 0.02 0.38 0.06 0.17 0.01 0.12 0.04 0.24 0.05 0.28 0.02 0.20 0.01 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 particular for ASL, ADSL, FAA, and FUM1. According to model iNX804 simulated flux distribution combined with the metabolic engineering results for ASL, ADSL, FAA and FUM1, ASL, and ADSL were confirmed to be the most effective pathways for producing fumaric acid. Moreover, ASL in the urea cycle and ADSL in the purine nucleotide cycle serve as a bridge between carbon metabolism and nitrogen metabolism, respectively. Therefore, such observations enhance the prevalent understanding of biosynthetic pathways for fumaric acid. The urea cycle implicates ornithine, citrulline, and arginine as participants in the synthesis of urea from aspartate and carbon dioxide (Krebs, 1982). This cycle has been identified in a variety of species, including E. coli and S. cerevisiae (Yu and Howell, 2000). The main functions of the urea cycle are the detoxification of waste nitrogen into excretable urea and the de novo biosynthesis of arginine (Brusilow et al., 2001). ASL catalyzes the reversible hydrolysis of argininosuccinate to arginine and fumaric acid, which is important for the detoxification of ammonia via the urea cycle and for arginine biosynthesis (Yu and Howell, 2000). Therefore, overexpression of ASL results in a significant increase in the fumaric acid titer, probably due to two aspects: (i) arginine, as the precursor for the synthesis of many biologically important compounds, including urea, polyamines, proline, and glutamate (Nagamani et al., 2012), plays roles in physiology and metabolism, including as an immediate precursor in protein synthesis, in the post-translational conjugation of arginine with the N-termini of proteins bearing Nterminal aspartate or glutamate, and as an allosteric activator of N-acetylglutamate synthase (Wu and Morris, 1998); (ii) fumaric acid, as an available TCA cycle intermediate, can tightly regulate cellular metabolism as an immediate precursor for protein synthesis, electron acceptor, and conserver of energy during electron transportation (Lambden and Guest, 1976). In most organisms, the purine biosynthetic pathway is nearly ubiquitous and results in the conversion of phosphoribosyl pyrophosphate to inosine 5’-monophosphate (IMP) (Kappock et al., 2000). ADSL plays a critical role in both cellular replication and metabolism via its action in the de novo purine biosynthetic pathway, in which it catalyzes two separate reactions, enabling it to participate in the addition of a nitrogen atom at two different positions in adenosine monophosphate (Toth and Yeates, 2000). These two separate reactions are the conversion of succinyladenosine monophosphate to adenosine monophosphate (AMP) and the conversion of succinylaminoimidazolecarboxamide ribonucleotide to aminoimidazolecarboxamide ribonucleotide (Spiegel et al., 2006). Both reactions catalyzed by ADSL involve the b-elimination of fumaric acid. Therefore, the fumaric acid titer is substantially increased by overexpressing ADSL, probably because the purine biosynthesis pathway not only provides the majority of purine nucleotides needed for cellular replication but also supplies amounts of available citric acid cycle intermediates. The purine nucleotide cycle aids in the tight regulation of cellular metabolism by controlling both the amounts of available citric acid cycle intermediates and the amount of free AMP (Toth and Yeates, 2000). In conclusion, the simultaneous overexpression of genes ASL and ADSL can effectively enhance the connection between carbon metabolism and nitrogen metabolism, improve the carbon flux distribution and increase fumaric acid production. The final concentration of fumaric acid increased to 5.62 g L1 in the strain T.G-ASL(H)-ADSL(L) by controlling the strength of ASL at a high level and ADSL at a low level, probably due to two aspects: (i) controlling ASL at a high level in the urea cycle is conducive to the detoxification of waste nitrogen into excretable urea and the de novo biosynthesis of fumaric acid and arginine (Brusilow et al., 2001); (ii) controlling ADSL at a low level in the purine biosynthesis pathway not only decreases the majority of purine nucleotides needed for cellular replication relatively but also increases amounts of available TCA cycle intermediates fumaric acid (Toth and Yeates, 2000). However, when FUM1 or FAA was overexpressed in T. glabrata, the pyruvate flux could not be more effectively redirected to fumaric acid compared to overexpression of ASL or ADSL. The reasons may be attributed to two factors: (i) FUM1, a key enzyme participating in the TCA cycle, catalyzes the reversible dehydration of malate to fumaric acid and exhibits higher affinity towards fumaric acid than towards malate (Xu et al., 2013a); (ii) FAA, participating in the catabolism of tyrosine, tryptophan, and phenylalanine in the cytosol, catalyzes the hydrolysis of fumarylacetoacetate to acetoacetate and fumaric acid and is thought to be genotoxic (Cassiman et al., 2009). Finally, further stepwise improvement was made by metabolic engineering based on overexpressing the SpMAE1 gene, thus achieving a higher fumaric acid titer of 8.83 g L1. However, the engineered strain T.G-ASL(H)-ADSL(L)SpMAE1 still produced 3.68 g L1 pyruvate, indicating that fumaric acid production can be further improved. With the aid of the ‘omic’ techniques, systems metabolic engineering emerges as a conceptual and technological framework to speed the modification of existing pathways for the optimal production of desired products, and has opened a novel avenue for engineering microorganisms to produce industrial products (Lee et al., 2012). For example, the bottleneck of producing fumaric acid with the urea cycle and the purine nucleotide cycle can be analyzed using transcriptome, proteome, and metabolomics profiling. After that, modular pathway engineering, systematic assembly, and optimization of metabolic modules is used for fine tuning existent pathways and balancing the metabolism of production hosts to debottleneck, debug, and improve cellular phenotypes (Juminaga et al., 2012). References Blazeck J, Alper H. 2010. Systems metabolic engineering: Genome-scale models and beyond. Biotechnol J 5(7):647–659. Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 165 Brusilow S, Horwich A, Scriver C, Beaudet A, Sly W, Valle D. 2001. The metabolic and molecular bases of inherited disease. 8th edn. McGraw-Hill. New York: 1909–1963. Canelas AB, Ras C, ten Pierick A, van Dam JC, Heijnen JJ, van Gulik WM. 2008. Leakage-free rapid quenching technique for yeast metabolomics. Metabolomics 4(3):226–239. Canelas AB, ten Pierick A, Ras C, Seifar RM, van Dam JC, van Gulik WM, Heijnen JJ. 2009. Quantitative evaluation of intracellular metabolite extraction techniques for yeast metabolomics. Anal Chem 81(17):7379– 7389. Carta FS, Soccol CR, Ramos LP, Fontana JD. 1999. Production of fumaric acid by fermentation of enzymatic hydrolysates derived from cassava bagasse. Bioresour Technol 68(1):23–28. Cassiman D, Zeevaert R, Holme E, Kvittingen EA, Jaeken J. 2009. A novel mutation causing mild, atypical fumarylacetoacetase deficiency (Tyrosinemia type I): A case report. Orphanet J Rare Dis 4:28. Chen X, Xu G, Xu N, Zou W, Zhu P, Liu L, Chen J. 2013. Metabolic engineering of Torulopsis glabrata for malate production. Metab Eng 19:10–16. Fu YQ, Xu Q, Li S, Chen Y, Huang H. 2010. Strain improvement of Rhizopus oryzae for over-production of fumaric acid by reducing ethanol synthesis pathway. Korean J Chem Eng 27(1):183–186. Gu S, Xu Q, Huang H, Li S. 2014. Alternative respiration and fumaric acid production of Rhizopus oryzae. Appl Microbiol Biotechnol 98(11):5145– 5152. Gusarova GA, Trejo HE, Dada LA, Briva A, Welch LC, Hamanaka RB, Mutlu GM, Chandel NS, Prakriya M, Sznajder JI. 2011. Hypoxia leads to Na, K-ATPase downregulation via Ca2þ release-activated Ca2þ channels and AMPK activation. Mol Cell Biol 31(17):3546–3556. Huang L, Wei PL, Zang R, Xu ZN, Cen PL. 2010. High-throughput screening of high-yield colonies of Rhizopus oryzae for enhanced production of fumaric acid. Ann Microbiol 60(2):287–292. Inoue H, Nojima H, Okayama H. 1990. High efficiency transformation of Escherichia coli with plasmids. Gene 96(1):23–28. Juminaga D, Baidoo EE, Redding-Johanson AM, Batth TS, Burd H, Mukhopadhyay A, Petzold CJ, Keasling JD. 2012. Modular engineering of L-tyrosine production in Escherichia coli. Appl Environ Microbiol 78(1):89–98. Kang SW, Lee H, Kim D, Lee D, Kim S, Chun GT, Lee J, Kim SW, Park C. 2010. Strain development and medium optimization for fumaric acid production. Biotechnol Bioproc E 15(5):761–769. Kappock TJ, Ealick SE, Stubbe J. 2000. Modular evolution of the purine biosynthetic pathway. Curr Opin Chem Biol 4(5):567–572. Krebs HA. 1982. The discovery of the ornithine cycle of urea synthesis. Trends Biochem Sci 7(2):76–78. Kvittingen E, Brodtkorb E. 1986. The pre-and post-natal diagnosis of tyrosinemia type I and the detection of the carrier state by assay of fumarylacetoacetase. Scand J Clin Lab Inv. 184:35. Lambden P, Guest J. 1976. Mutants of Escherichia coli K12 unable to use fumarate as an anaerobic electron acceptor. J Gen Microbiol 97(2): 145–160. Lee JW, Na D, Park JM, Lee J, Choi S, Lee SY. 2012. Systems metabolic engineering of microorganisms for natural and non-natural chemicals. Nat Chem Biol 8(6):536–546. Lee P, Colman RF. 2007. Expression, purification, and characterization of stable, recombinant human adenylosuccinate lyase. Protein Expr Purif 51(2):227–234. Lee TT, Worby C, Dixon JE, Colman RF. 1997. Identification of His(141) in the active site of Bacillus subtilis adenylosuccinate lyase by affinity labeling with 6-(4-bromo-2,3-dioxobutyl)thioadenosine 5’-monophosphate. J Biol Chem 272(1):458–465. Liu LM, Li Y, Li HZ, Chen J. 2004a. Manipulating the pyruvate dehydrogenase bypass of a multi-vitamin auxotrophic yeast Torulopsis glabrata enhanced pyruvate production. Lett Appl Microbiol 39(2):199– 206. Liu LM, Li Y, Li HZ, Chen J. 2004b. Manipulating the pyruvate dehydrogenase bypass of a multi-vitamin auxotrophic yeast Torulopsis 166 Biotechnology and Bioengineering, Vol. 112, No. 1, January, 2015 glabrata enhanced pyruvate production. Lett Appl Microbiol 39(2):199– 206. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the Folin phenol reagent. J Biol Chem 193(1):265–275. Mizutani H, Kunishima N. 2007. Purification, crystallization and preliminary X-ray analysis of the fumarylacetoacetase family member TTHA0809 from Thermus thermophilus HB8. Acta Cryst F63:792–794. Nagamani SC, Erez A, Lee B. 2012. Argininosuccinate lyase deficiency. Genet Med 14(5):501–507. Riscaldati E, Moresi M, Federici F, Petruccioli M. 2000. Direct ammonium fumarate production by Rhizopus arrhizus under phosphorous limitation. Biotechnol Lett 22(13):1043–1047. Roa Engel C. 2010. Integration of fermentation and cooling crystallisation to produce organic acids. Delft University of Technology: 87–108. Roa Engel CA, Straathof AJ, Zijlmans TW, van Gulik WM, van der Wielen LA. 2008. Fumaric acid production by fermentation. Appl Microbiol Biotechnol 78(3):379–389. Sadeh A, Baran D, Volokh M, Aharoni A. 2012. Conserved motifs in the Msn2-activating domain are important for Msn2-mediated yeast stress response. J Cell Sci 125:3333–3342. Shih VE, Littlefield JW, Moser HW. 1969. Argininosuccinase deficiency in fibroblasts cultured from patients with argininosuccinic aciduria. Biochem Genet 3(1):81–83. Song CW, Kim DI, Choi S, Jang JW, Lee SY. 2013. Metabolic engineering of Escherichia coli for the production of fumaric acid. Biotechnol Bioeng 110:2025–2034. Spiegel EK, Colman RF, Patterson D. 2006. Adenylosuccinate lyase deficiency. Mol Genet Metab 89(1–2):19–31. Toth EA, Yeates TO. 2000. The structure of adenylosuccinate lyase, an enzyme with dual activity in the de novo purine biosynthetic pathway. Structure 8(2):163–174. Trevisson E, Burlina A, Doimo M, Pertegato V, Casarin A, Cesaro L, Navas P, Basso G, Sartori G, Salviati L. 2009. Functional complementation in yeast allows molecular characterization of missense argininosuccinate lyase mutations. J Biol Chem 284(42):28926–28934. Wang G, Huang D, Qi H, Wen J, Jia X, Chen Y. 2013. Rational medium optimization based on comparative metabolic profiling analysis to improve fumaric acid production. Bioresour Technol 137:1–8. Werpy T, Petersen G. 2004. Top value added chemicals from biomass: Volume I Results of screening for potential candidates from sugars and synthesis gas. University of Pennsylvania Law Review 54(3):477. Wu G, Morris SM Jr. 1998. Arginine metabolism: Nitric oxide and beyond. Biochem J 336(Pt1):1–17. Xu G, Chen X, Liu L, Jiang L. 2013a. Fumaric acid production in Saccharomyces cerevisiae by simultaneous use of oxidative and reductive routes. Bioresour Technol 148:91–96. Xu G, Liu L, Chen J. 2012a. Reconstruction of cytosolic fumaric acid biosynthetic pathways in Saccharomyces cerevisiae. Microb Cell Fact 11:24. Xu G, Zou W, Chen X, Xu N, Liu L, Chen J. 2012b. Fumaric acid production in Saccharomyces cerevisiae by in silico aided metabolic engineering. PLoS One 7(12):52086. Xu M, Rao Z, Xu H, Lan C, Dou W, Zhang X, Xu H, Jin J, Xu Z. 2011. Enhanced production of L-arginine by expression of Vitreoscilla hemoglobin using a novel expression system in Corynebacterium crenatum. Appl Biochem Biotechnol 163(6):707–719. Xu N, Liu L, Zou W, Liu J, Hua Q, Chen J. 2013b. Reconstruction and analysis of the genome-scale metabolic network of Candida glabrata. Mol Biosyst 9(2):205–216. Xu Q, Li S, Huang H, Wen J. 2012c. Key technologies for the industrial production of fumaric acid by fermentation. Biotechnol Adv 30(6):1685–1696. Xu S, Zhou J, Qin Y, Liu L, Chen J. 2010. Water-forming NADH oxidase protects Torulopsis glabrata against hyperosmotic stress. Yeast 27(4):207– 216. Yu B, Howell PL. 2000. Intragenic complementation and the structure and function of argininosuccinate lyase. Cell Mol Life Sci 57(11):1637–1651. Zelle RM, de Hulster E, van Winden WA, de Waard P, Dijkema C, Winkler AA, Geertman JM, van Dijken JP, Pronk JT, van Maris AJ. 2008. Malic acid production by Saccharomyces cerevisiae: Engineering of pyruvate carboxylation, oxaloacetate reduction, and malate export. Appl Environ Microbiol 74(9):2766–2777. Zhang B, Skory CD, Yang ST. 2012. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing pyc and pepc genes on fumaric acid biosynthesis from glucose. Metab Eng 14(5):512–520. Zhou J, Dong Z, Liu L, Du G, Chen J. 2009. A reusable method for construction of non-marker large fragment deletion yeast auxotroph strains: A practice in Torulopsis glabrata. J Microbiol Methods 76(1): 70–74. Zhou Y, Du J, Tsao GT. 2002. Comparison of fumaric acid production by Rhizopus oryzae using different neutralizing agents. Bioprocess Biosyst Eng 25(3):179–181. Zhou Y, Nie K, Zhang X, Liu S, Wang M, Deng L, Wang F, Tan T. 2014. Production of fumaric acid from biodiesel-derived crude glycerol by Rhizopus arrhizus. Bioresour Technol 163C:48–53. Chen et al.: Fumaric Acid Production by Torulopsis glabrata Biotechnology and Bioengineering 167